

The disease-causing mutations in the carboxyl terminus of the cone cyclic nucleotide-gated channel CNGA3 subunit alter the local secondary structure and interfere with the channel active conformational change
- PMID: 20088482
- PMCID: PMC2829136
- DOI: 10.1021/bi901960u
The disease-causing mutations in the carboxyl terminus of the cone cyclic nucleotide-gated channel CNGA3 subunit alter the local secondary structure and interfere with the channel active conformational change
Abstract
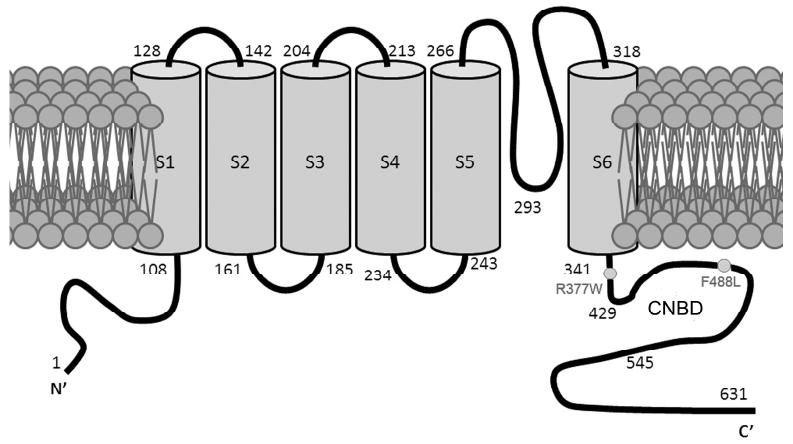
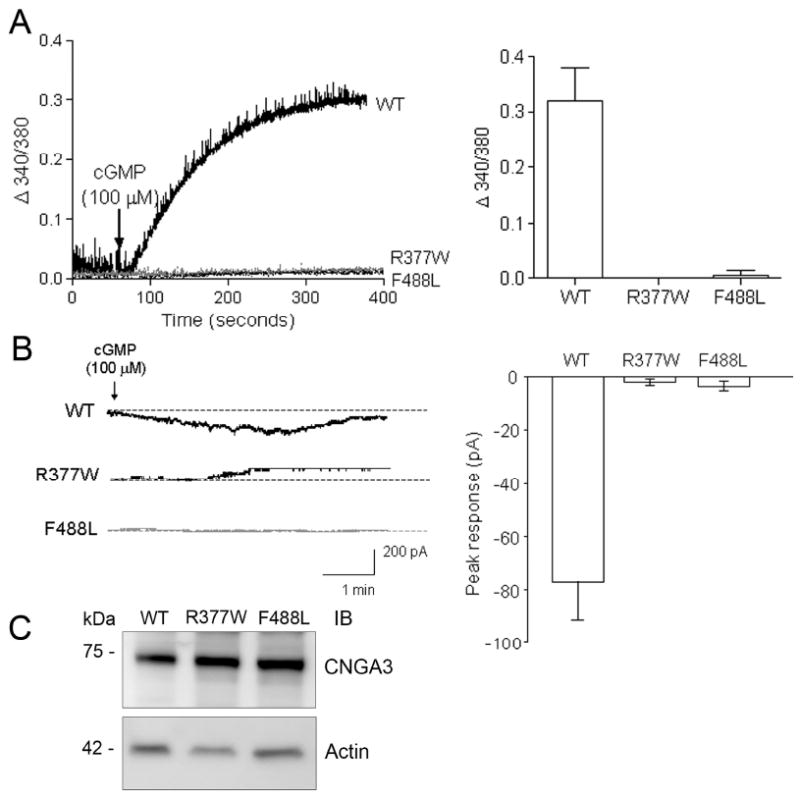
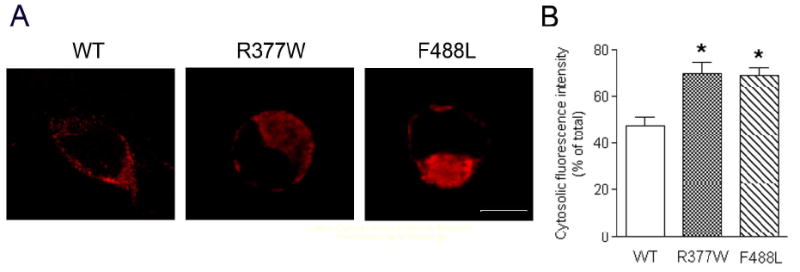
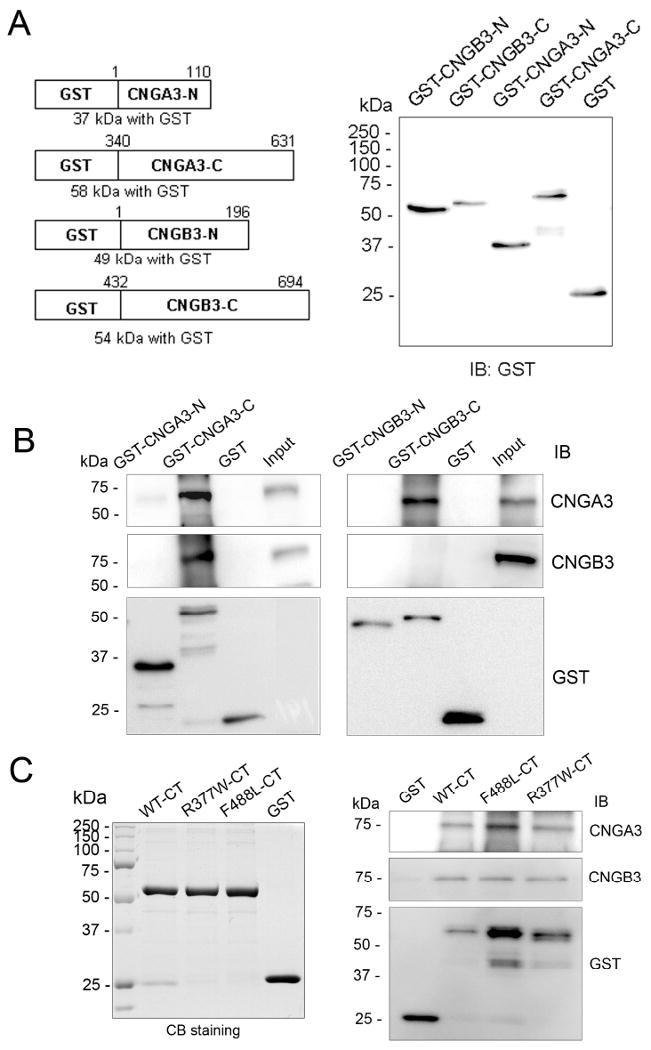
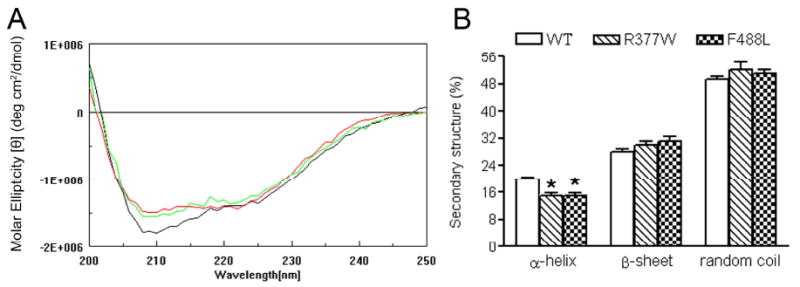
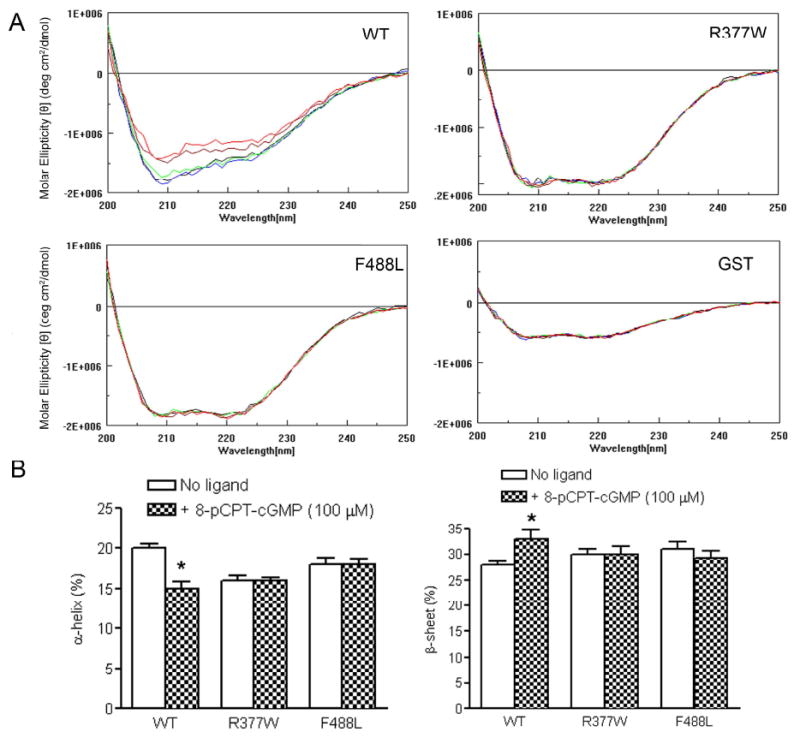
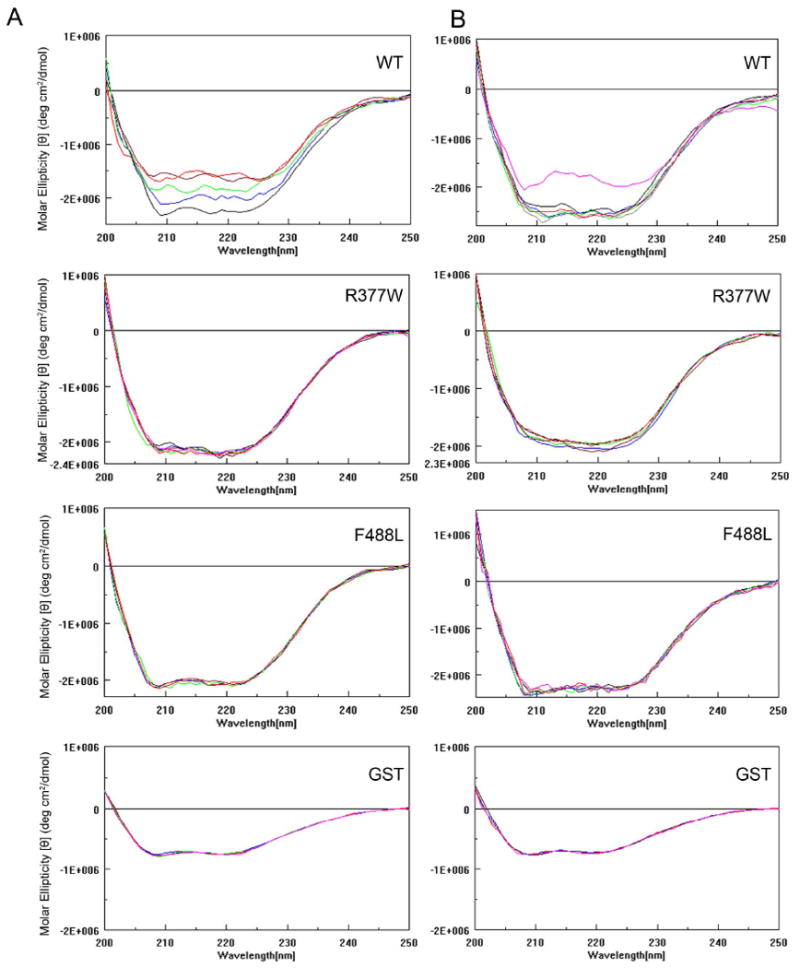
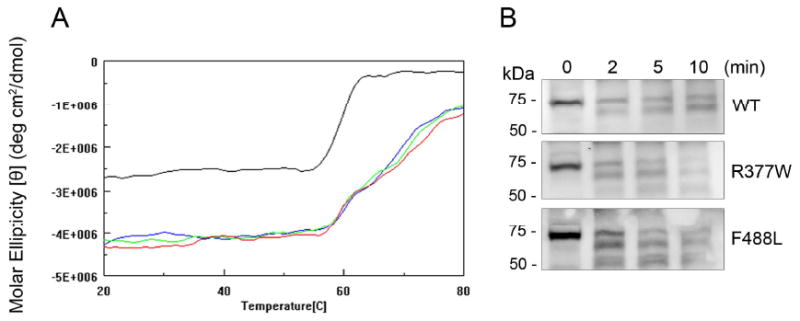
The cone photoreceptor cyclic nucleotide-gated (CNG) channel plays a pivotal role in phototransducton. Mutations in the channel subunits are associated with achromatopsia and progressive cone dystrophy in humans. More than 50 mutations have been identified in the channel CNGA3 subunit, with 50% of them located in the carboxyl (C) terminus. This study investigates the defects of the two frequently occurring mutations, R377W and F488L, in the C-terminus of CNGA3. Ratiometric measurement of the intracellular Ca(2+) concentration and electrophysiological recordings showed the loss of functional activity of the mutant channels in an HEK293 heterologous expression system. Immunofluorescence labeling revealed an apparent cytosolic aggregation of the mutant channels compared to the wild type (WT). The R377W and F488L mutants, expressed and purified from Escherichia coli as glutathione S-transferase (GST) fused to the CNGA3 C-terminal domain, showed no negative effects on interactions with the channel subunits. Circular dichroism spectrum analyses were performed to examine the structural impact of the mutations. Although the R377W and F488L C-termini mutants retained stable, folded structures, the secondary structures of both mutants differed from the WT protein. Furthermore, the WT C-terminus exhibited a significant decrease in alpha-helical content in response to the channel ligands, while this allosteric transition was diminished in the two mutants. This is the first study showing the structural impact of the disease-causing mutations in the cone CNG channel subunit. The observed alterations in the local secondary structure and active conformational change may confer an adverse effect on the channel's activity and cellular processing.
Figures
Similar articles
-
Molecular pathogenesis of achromatopsia associated with mutations in the cone cyclic nucleotide-gated channel CNGA3 subunit.Adv Exp Med Biol. 2010;664:245-53. doi: 10.1007/978-1-4419-1399-9_28. Adv Exp Med Biol. 2010. PMID: 20238023 Free PMC article.
-
CNGA3 achromatopsia-associated mutation potentiates the phosphoinositide sensitivity of cone photoreceptor CNG channels by altering intersubunit interactions.Am J Physiol Cell Physiol. 2013 Jul 15;305(2):C147-59. doi: 10.1152/ajpcell.00037.2013. Epub 2013 Apr 3. Am J Physiol Cell Physiol. 2013. PMID: 23552282 Free PMC article.
-
Achromatopsia-associated mutation in the human cone photoreceptor cyclic nucleotide-gated channel CNGB3 subunit alters the ligand sensitivity and pore properties of heteromeric channels.J Biol Chem. 2003 Sep 5;278(36):34533-40. doi: 10.1074/jbc.M305102200. Epub 2003 Jun 18. J Biol Chem. 2003. PMID: 12815043
-
Retinal Cyclic Nucleotide-Gated Channel Regulation by Calmodulin.Int J Mol Sci. 2022 Nov 16;23(22):14143. doi: 10.3390/ijms232214143. Int J Mol Sci. 2022. PMID: 36430626 Free PMC article. Review.
-
Comprehensive variant spectrum of the CNGA3 gene in patients affected by achromatopsia.Hum Mutat. 2022 Jul;43(7):832-858. doi: 10.1002/humu.24371. Epub 2022 Apr 14. Hum Mutat. 2022. PMID: 35332618 Review.
Cited by
-
Achromatopsia: Genetics and Gene Therapy.Mol Diagn Ther. 2022 Jan;26(1):51-59. doi: 10.1007/s40291-021-00565-z. Epub 2021 Dec 3. Mol Diagn Ther. 2022. PMID: 34860352 Free PMC article. Review.
-
The B3 Subunit of the Cone Cyclic Nucleotide-gated Channel Regulates the Light Responses of Cones and Contributes to the Channel Structural Flexibility.J Biol Chem. 2016 Apr 15;291(16):8721-34. doi: 10.1074/jbc.M115.696138. Epub 2016 Feb 18. J Biol Chem. 2016. PMID: 26893377 Free PMC article.
-
Early-onset, slow progression of cone photoreceptor dysfunction and degeneration in CNG channel subunit CNGB3 deficiency.Invest Ophthalmol Vis Sci. 2011 Jun 1;52(6):3557-66. doi: 10.1167/iovs.10-6358. Invest Ophthalmol Vis Sci. 2011. PMID: 21273547 Free PMC article.
-
Defective trafficking of cone photoreceptor CNG channels induces the unfolded protein response and ER-stress-associated cell death.Biochem J. 2012 Jan 15;441(2):685-96. doi: 10.1042/BJ20111004. Biochem J. 2012. PMID: 21992067 Free PMC article.
-
Biochemical characterization of cone cyclic nucleotide-gated (CNG) channel using the infrared fluorescence detection system.Adv Exp Med Biol. 2012;723:769-75. doi: 10.1007/978-1-4614-0631-0_98. Adv Exp Med Biol. 2012. PMID: 22183405 Free PMC article.
References
-
- Zagotta WN, Siegelbaum SA. Structure and function of cyclic nucleotide-gated channels. Annu Rev Neurosci. 1996;19:235–263. - PubMed
-
- Kaupp UB, Seifert R. Cyclic nucleotide-gated ion channels. Physiol Rev. 2002;82:769–824. - PubMed
-
- Weitz D, Ficek N, Kremmer E, Bauer PJ, Kaupp UB. Subunit stoichiometry of the CNG channel of rod photoreceptors. Neuron. 2002;36:881–889. - PubMed
-
- Peng C, Rich ED, Varnum MD. Subunit configuration of heteromeric cone cyclic nucleotide-gated channels. Neuron. 2004;42:401–410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
