

Inactivation of Lactobacillus leichmannii ribonucleotide reductase by 2',2'-difluoro-2'-deoxycytidine 5'-triphosphate: adenosylcobalamin destruction and formation of a nucleotide-based radical
- PMID: 20088568
- PMCID: PMC2867335
- DOI: 10.1021/bi9021318
Inactivation of Lactobacillus leichmannii ribonucleotide reductase by 2',2'-difluoro-2'-deoxycytidine 5'-triphosphate: adenosylcobalamin destruction and formation of a nucleotide-based radical
Abstract
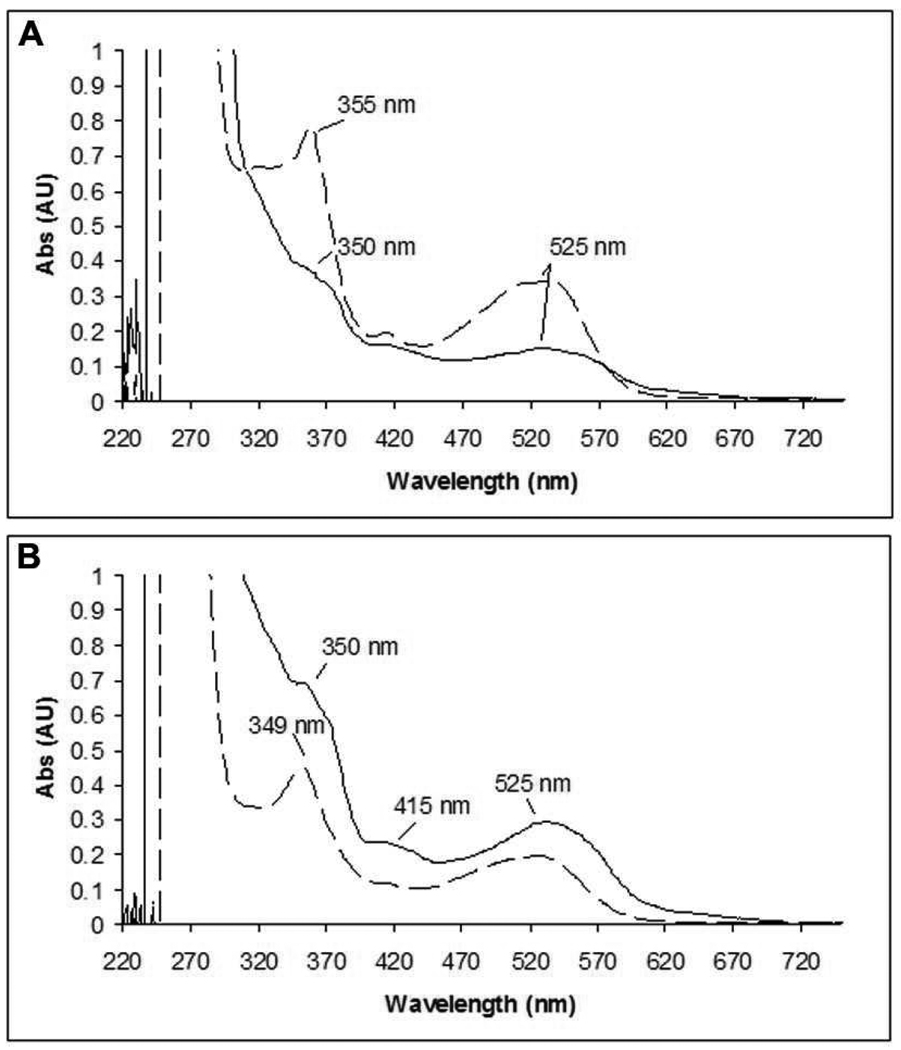
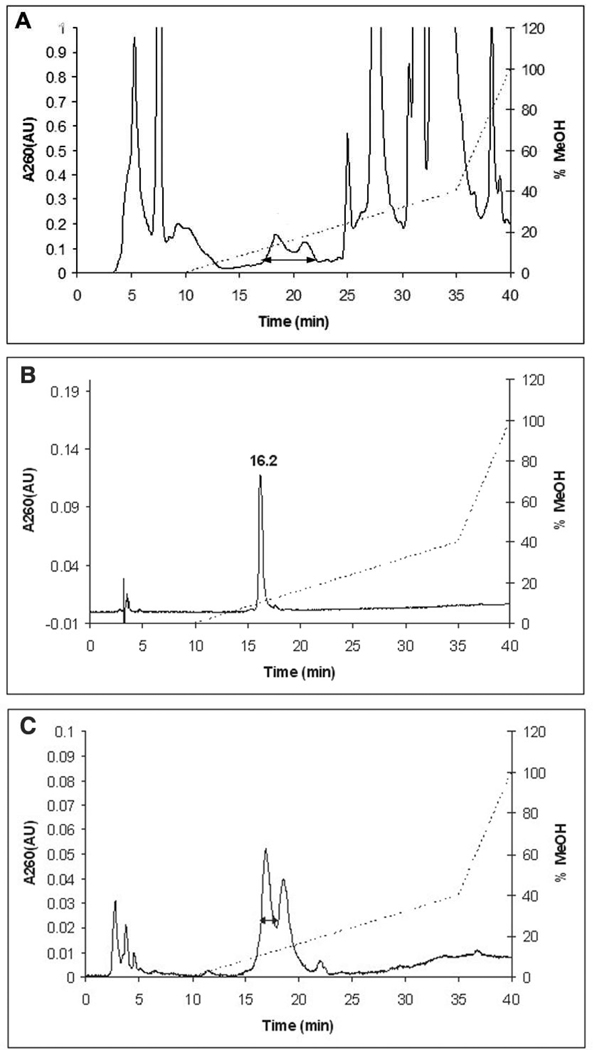
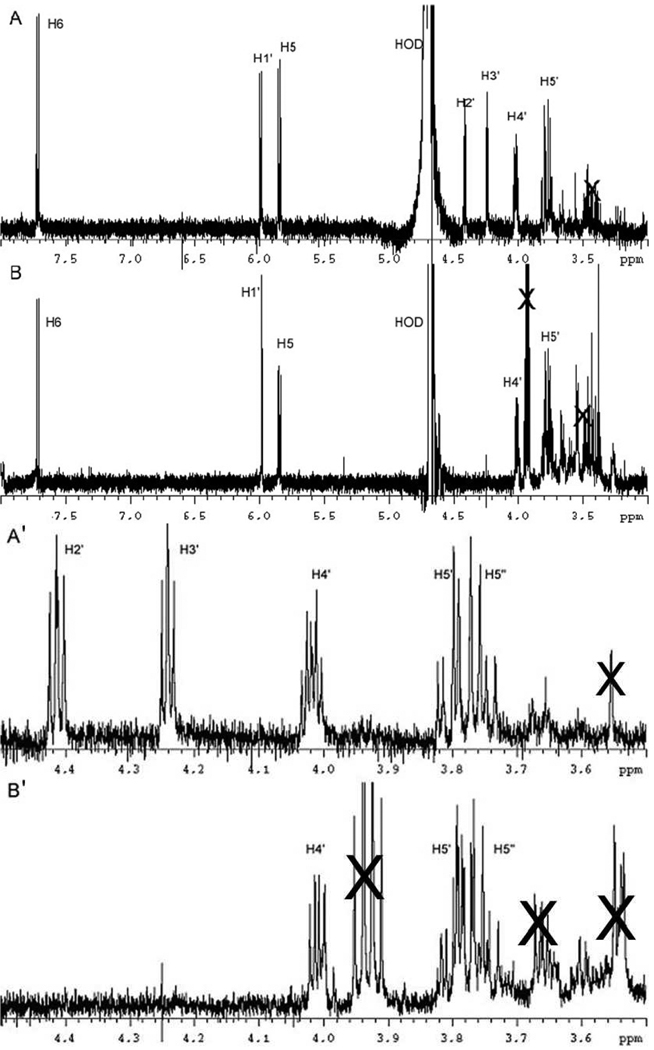
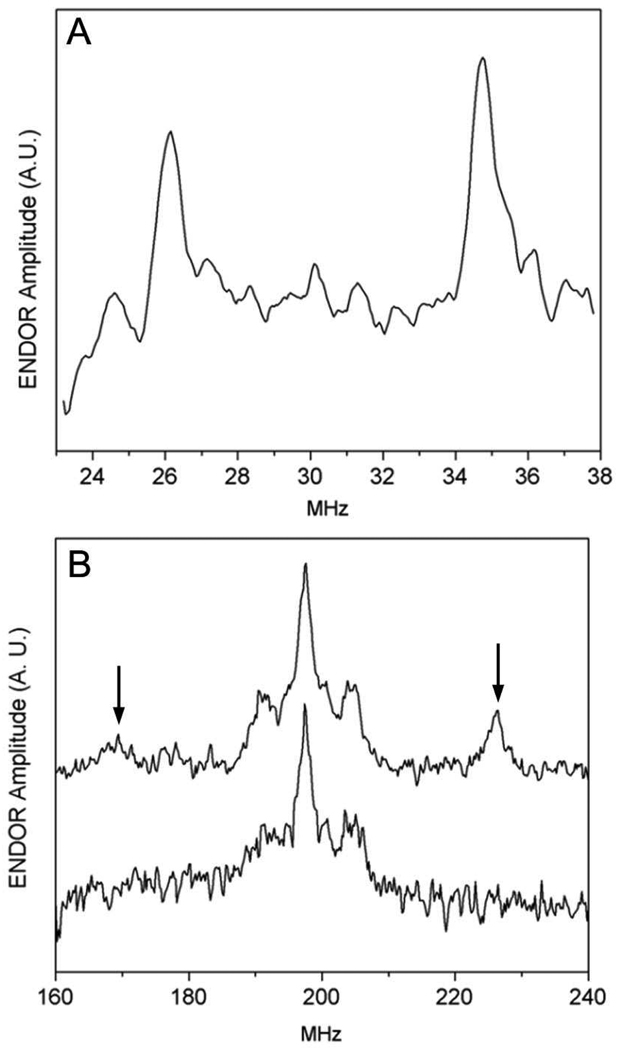
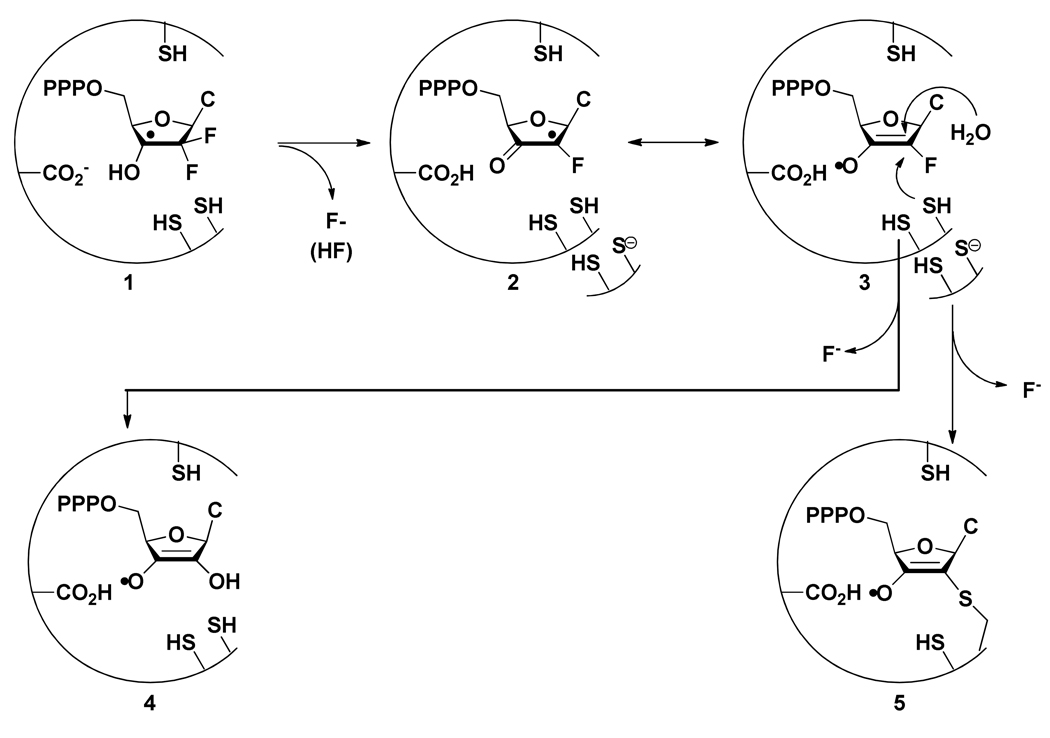
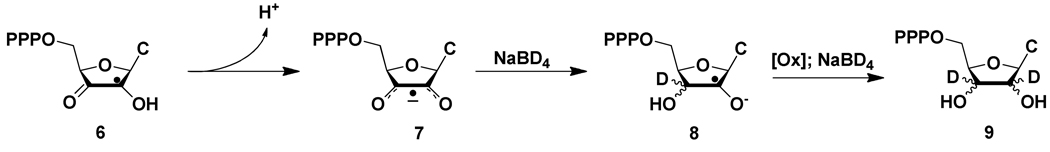
Ribonucleotide reductase (RNR, 76 kDa) from Lactobacillus leichmannii is a class II RNR that requires adenosylcobalamin (AdoCbl) as a cofactor. It catalyzes the conversion of nucleoside triphosphates to deoxynucleotides and is 100% inactivated by 1 equiv of 2',2'-difluoro-2'-deoxycytidine 5'-triphosphate (F(2)CTP) in <2 min. Sephadex G-50 chromatography of the inactivation reaction mixture for 2 min revealed that 0.47 equiv of a sugar moiety is covalently bound to RNR and 0.25 equiv of a cobalt(III) corrin is tightly associated, likely through a covalent interaction with C(419) (Co-S) in the active site of RNR [Lohman, G. J. S., and Stubbe, J. (2010) Biochemistry 49, DOI: 10.1021/bi902132u ]. After 1 h, a similar experiment revealed 0.45 equiv of the Co-S adduct associated with the protein. Thus, at least two pathways are associated with RNR inactivation: one associated with alkylation by the sugar of F(2)CTP and the second with AdoCbl destruction. To determine the fate of [1'-(3)H]F(2)CTP in the latter pathway, the reaction mixture at 2 min was reduced with NaBH(4) (NaB(2)H(4)) and the protein separated from the small molecules using a centrifugation device. The small molecules were dephosphorylated and analyzed by HPLC to reveal 0.25 equiv of a stereoisomer of cytidine, characterized by mass spectrometry and NMR spectroscopy, indicating the trapped nucleotide had lost both of its fluorides and gained an oxygen. High-field ENDOR studies with [1'-(2)H]F(2)CTP from the reaction quenched at 30 s revealed a radical that is nucleotide-based. The relationship between this radical and the trapped cytidine analogue provides insight into the nonalkylative pathway for RNR inactivation relative to the alkylative pathway.
Figures
Similar articles
-
Inactivation of Lactobacillus leichmannii ribonucleotide reductase by 2',2'-difluoro-2'-deoxycytidine 5'-triphosphate: covalent modification.Biochemistry. 2010 Feb 23;49(7):1404-17. doi: 10.1021/bi902132u. Biochemistry. 2010. PMID: 20088569 Free PMC article.
-
Gemcitabine 5'-triphosphate is a stoichiometric mechanism-based inhibitor of Lactobacillus leichmannii ribonucleoside triphosphate reductase: evidence for thiyl radical-mediated nucleotide radical formation.Biochemistry. 1998 Apr 21;37(16):5528-35. doi: 10.1021/bi972934e. Biochemistry. 1998. PMID: 9548936
-
Studies on the catalysis of carbon-cobalt bond homolysis by ribonucleoside triphosphate reductase: evidence for concerted carbon-cobalt bond homolysis and thiyl radical formation.Biochemistry. 1999 Jan 26;38(4):1221-33. doi: 10.1021/bi981885i. Biochemistry. 1999. PMID: 9930982
-
The function of adenosylcobalamin in the mechanism of ribonucleoside triphosphate reductase from Lactobacillus leichmannii.Curr Opin Chem Biol. 1998 Oct;2(5):650-5. doi: 10.1016/s1367-5931(98)80097-5. Curr Opin Chem Biol. 1998. PMID: 9818192 Review.
-
The enzymatic activation of coenzyme B12.Dalton Trans. 2006 Mar 7;(9):1123-33. doi: 10.1039/b517599m. Epub 2006 Feb 7. Dalton Trans. 2006. PMID: 16482346 Review.
Cited by
-
Inhibitors of the Cancer Target Ribonucleotide Reductase, Past and Present.Biomolecules. 2022 Jun 10;12(6):815. doi: 10.3390/biom12060815. Biomolecules. 2022. PMID: 35740940 Free PMC article. Review.
-
A chemically competent thiosulfuranyl radical on the Escherichia coli class III ribonucleotide reductase.J Am Chem Soc. 2014 Jun 25;136(25):9001-13. doi: 10.1021/ja5030194. Epub 2014 Jun 17. J Am Chem Soc. 2014. PMID: 24827372 Free PMC article.
-
Insight into the mechanism of inactivation of ribonucleotide reductase by gemcitabine 5'-diphosphate in the presence or absence of reductant.Biochemistry. 2009 Dec 15;48(49):11622-9. doi: 10.1021/bi901590q. Biochemistry. 2009. PMID: 19899770 Free PMC article.
-
Model Substrate/Inactivation Reactions for MoaA and Ribonucleotide Reductases: Loss of Bromo, Chloro, or Tosylate Groups from C2 of 1,5-Dideoxyhomoribofuranoses upon Generation of an α-Oxy Radical at C3.Molecules. 2020 May 29;25(11):2539. doi: 10.3390/molecules25112539. Molecules. 2020. PMID: 32486052 Free PMC article.
-
Modulation of the ribonucleotide reductase M1-gemcitabine interaction in vivo by N-ethylmaleimide.Biochem Biophys Res Commun. 2011 Sep 23;413(2):383-8. doi: 10.1016/j.bbrc.2011.08.111. Epub 2011 Aug 27. Biochem Biophys Res Commun. 2011. PMID: 21893046 Free PMC article.
References
-
- Stubbe J, van der Donk WA. Ribonucleotide reductases: radical enzymes with suicidal tendencies. Chem. Biol. 1995;2:793–801. - PubMed
-
- Stubbe J, van der Donk WA. Protein radicals in enzyme catalysis. Chem. Rev. 1998;98:705–762. - PubMed
-
- Licht S, Stubbe J. Mechanistic investigations of ribonucleotide reductases. In: Barton SD, Nakanishi K, Meth-Cohn O, Poulter CD, editors. Comprehensive Natural Products Chemistry. New York: Elsevier Science; 1999. pp. 163–203.
-
- Eklund H, Uhlin U, Färnegårdh M, Logan DT, Nordlund P. Structure and function of the radical enzyme ribonucleotide reductase. Prog. Biophys. Mol. Biol. 2001;77:177–268. - PubMed
-
- Nordlund P, Reichard P. Ribonucleotide reductases. Annu. Rev. Biochem. 2006;75:681–706. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
