

Release of SF3 from the intron branchpoint activates the first step of pre-mRNA splicing
- PMID: 20089683
- PMCID: PMC2822917
- DOI: 10.1261/rna.2030510
Release of SF3 from the intron branchpoint activates the first step of pre-mRNA splicing
Abstract
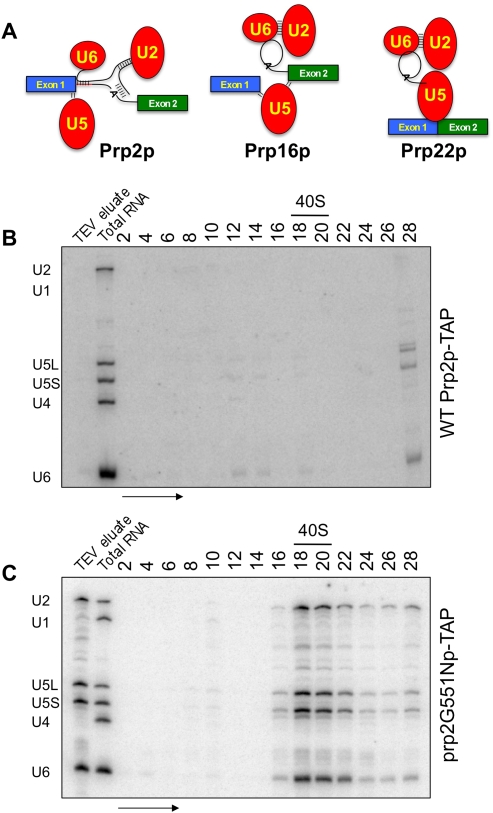
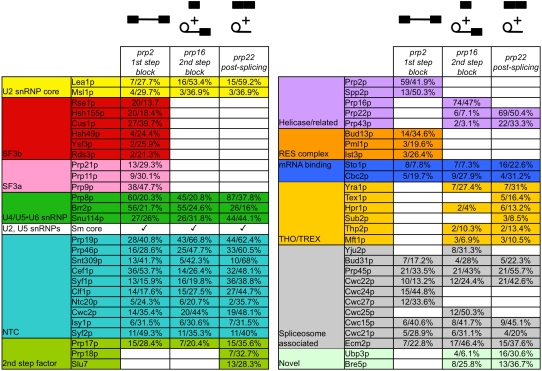
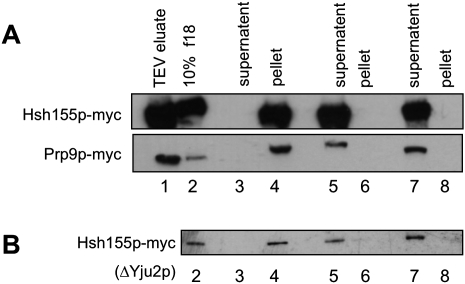
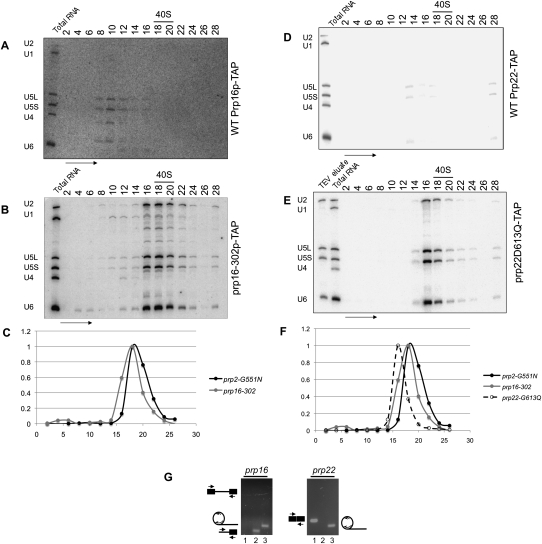
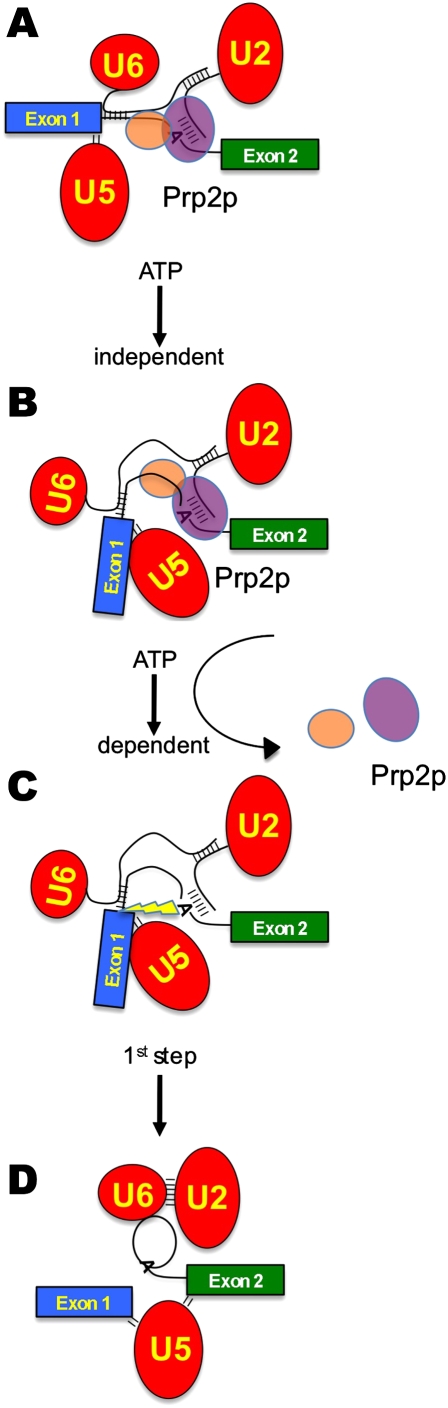
Eukaryotic pre-mRNA splicing is a complex process requiring the precise timing and action of >100 trans-acting factors. It has been known for some time that the two steps of splicing chemistry require three DEAH-box RNA helicase-like proteins; however, their mechanism of action at these steps has remained elusive. Spliceosomes arrested in vivo at the three helicase checkpoints were purified, and first step-arrested spliceosomes were functionally characterized. We show that the first step of splicing requires a novel ATP-independent conformational change. Prp2p then catalyzes an ATP-dependent rearrangement displacing the SF3a and SF3b complexes from the branchpoint within the spliceosome. We propose a model in which SF3 prevents premature nucleophilic attack of the chemically reactive hydroxyl of the branchpoint adenosine prior to the first transesterification. When the spliceosome attains the proper conformation and upon the function of Prp2p, SF3 is displaced from the branchpoint allowing first step chemistry to occur.
Figures

References
-
- Baker-Brachmann C, Davies A, Cost GJ, Caputo E, Li J, Hieter P, Boeke JD. Designer deletion strains derived from Saccharomyces cerevisiae S288C: A useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast. 1998;14:115–132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases