

R2R3-NaMYB8 regulates the accumulation of phenylpropanoid-polyamine conjugates, which are essential for local and systemic defense against insect herbivores in Nicotiana attenuata
- PMID: 20089770
- PMCID: PMC2832263
- DOI: 10.1104/pp.109.151738
R2R3-NaMYB8 regulates the accumulation of phenylpropanoid-polyamine conjugates, which are essential for local and systemic defense against insect herbivores in Nicotiana attenuata
Abstract
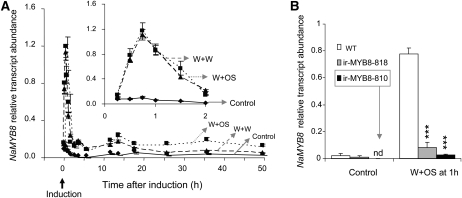
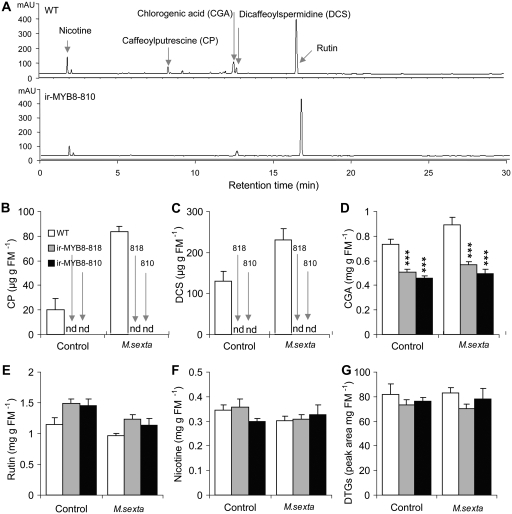
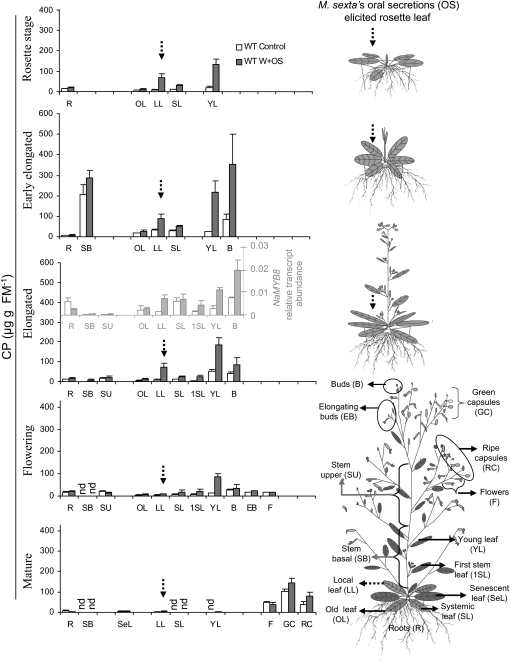
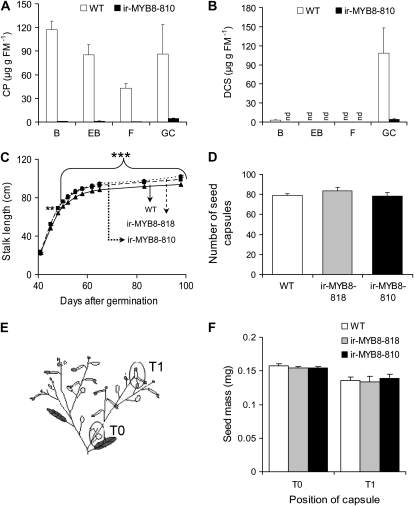
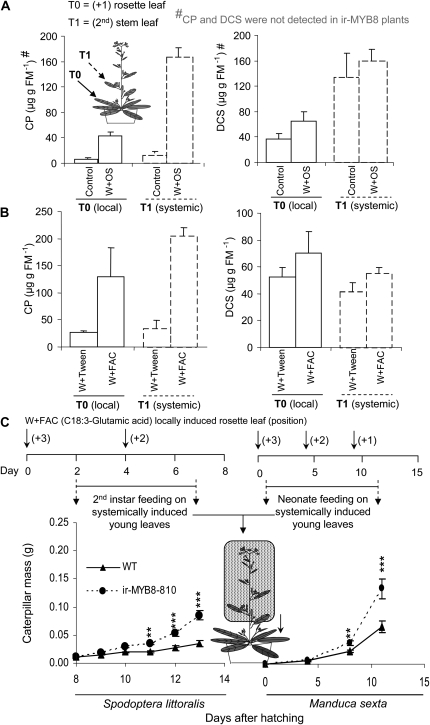
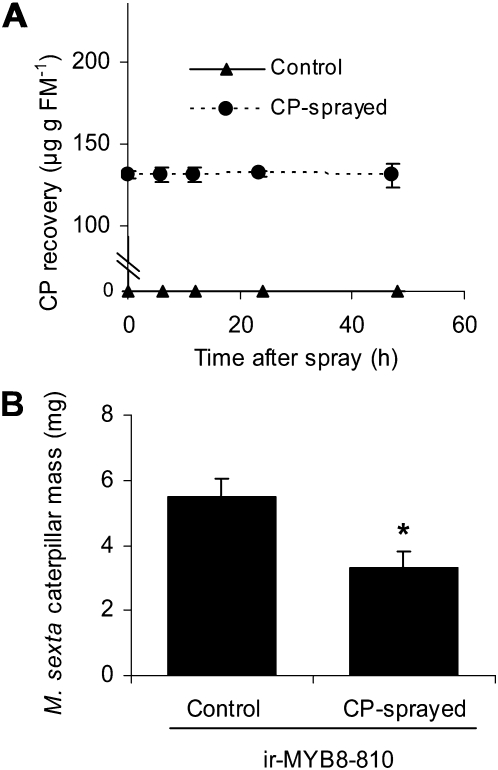
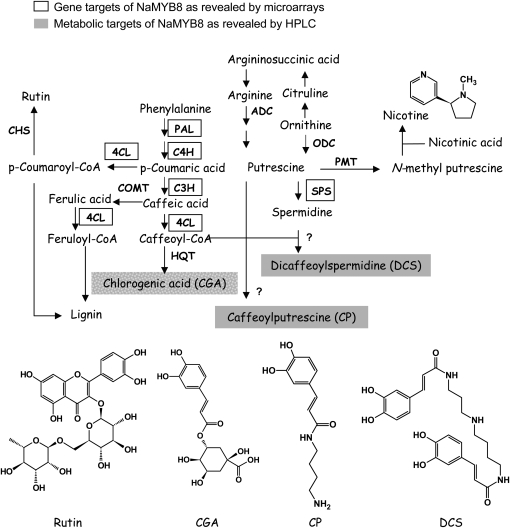
Although phenylpropanoid-polyamine conjugates (PPCs) occur ubiquitously in plants, their biological roles remain largely unexplored. The two major PPCs of Nicotiana attenuata plants, caffeoylputrescine (CP) and dicaffeoylspermidine, increase dramatically in local and systemic tissues after herbivore attack and simulations thereof. We identified NaMYB8, a homolog of NtMYBJS1, which in BY-2 cells regulates PPC biosynthesis, and silenced its expression by RNA interference in N. attenuata (ir-MYB8), to understand the ecological role(s) of PPCs. The regulatory role of NaMYB8 in PPC biosynthesis was validated by a microarray analysis, which revealed that transcripts of several key biosynthetic genes in shikimate and polyamine metabolism accumulated in a NaMYB8-dependent manner. Wild-type N. attenuata plants typically contain high levels of PPCs in their reproductive tissues; however, NaMYB8-silenced plants that completely lacked CP and dicaffeoylspermidine showed no changes in reproductive parameters of the plants. In contrast, a defensive role for PPCs was clear; both specialist (Manduca sexta) and generalist (Spodoptera littoralis) caterpillars feeding on systemically preinduced young stem leaves performed significantly better on ir-MYB8 plants lacking PPCs compared with wild-type plants expressing high levels of PPCs. Moreover, the growth of M. sexta caterpillars was significantly reduced when neonates were fed ir-MYB8 leaves sprayed with synthetic CP, corroborating the role of PPCs as direct plant defense. The spatiotemporal accumulation and function of PPCs in N. attenuata are consistent with the predictions of the optimal defense theory: plants preferentially protect their most fitness-enhancing and vulnerable parts, young tissues and reproductive organs, to maximize their fitness.
Figures
References
-
- Balint R, Cooper G, Staebell M, Filner P. (1987) N-Caffeoyl-4-amino-n-butyric acid, a new flower-specific metabolite in cultured tobacco cells and tobacco plants. J Biol Chem 262: 11026–11031 - PubMed
-
- Baumann K, Perez-Rodriguez M, Bradley D, Venail J, Bailey P, Jin H, Koes R, Roberts K, Martin C. (2007) Control of cell and petal morphogenesis by R2R3 MYB transcription factors. Development 134: 1691–1701 - PubMed
-
- Boege K, Dirzo R, Siemens D, Brown P. (2007) Ontogenetic switches from plant resistance to tolerance: minimizing costs with age?. Ecol Lett 10: 177–187 - PubMed
-
- Boege K, Marquis RJ. (2005) Facing herbivory as you grow up: the ontogeny of resistance in plants. Trends Ecol Evol 20: 441–448 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
