

Neural integration of information specifying human structure from form, motion, and depth
- PMID: 20089892
- PMCID: PMC2819284
- DOI: 10.1523/JNEUROSCI.3116-09.2010
Neural integration of information specifying human structure from form, motion, and depth
Abstract
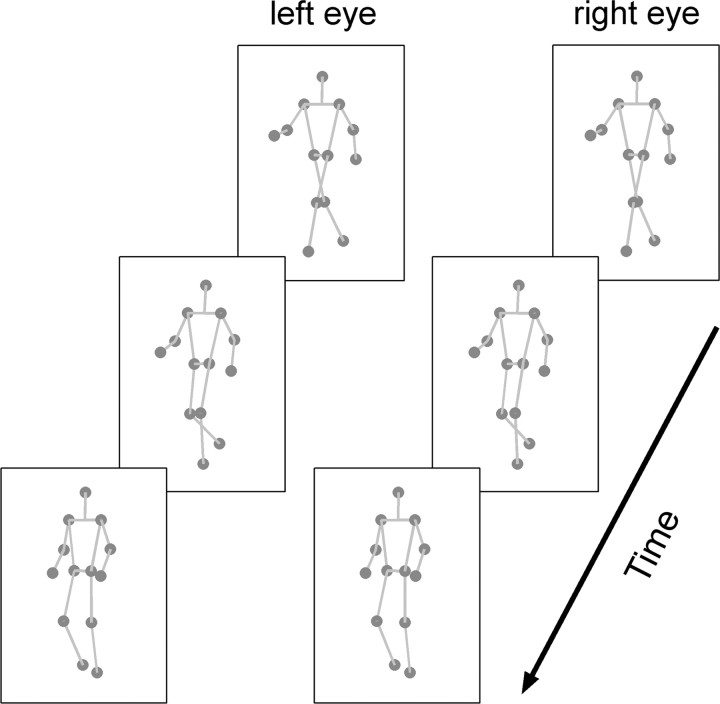
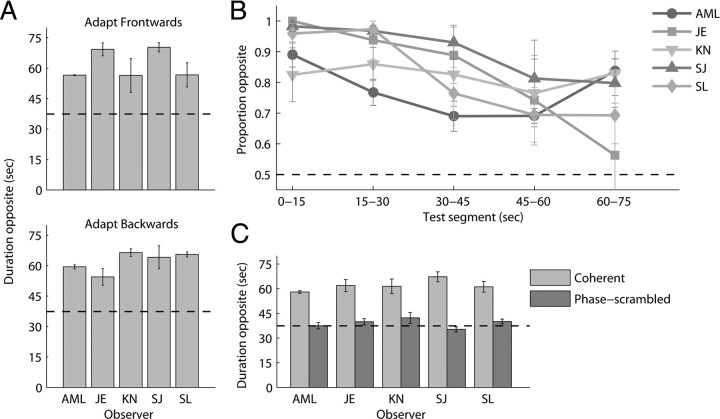
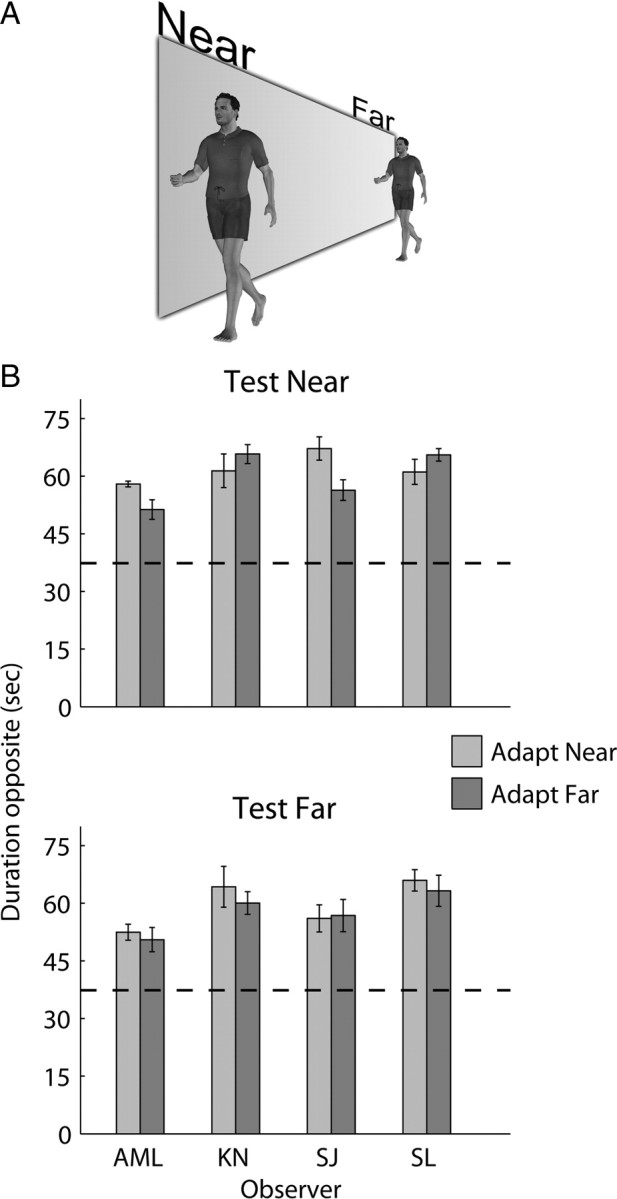
Recent computational models of biological motion perception operate on ambiguous two-dimensional representations of the body (e.g., snapshots, posture templates) and contain no explicit means for disambiguating the three-dimensional orientation of a perceived human figure. Are there neural mechanisms in the visual system that represent a moving human figure's orientation in three dimensions? To isolate and characterize the neural mechanisms mediating perception of biological motion, we used an adaptation paradigm together with bistable point-light (PL) animations whose perceived direction of heading fluctuates over time. After exposure to a PL walker with a particular stereoscopically defined heading direction, observers experienced a consistent aftereffect: a bistable PL walker, which could be perceived in the adapted orientation or reversed in depth, was perceived predominantly reversed in depth. A phase-scrambled adaptor produced no aftereffect, yet when adapting and test walkers differed in size or appeared on opposite sides of fixation aftereffects did occur. Thus, this heading direction aftereffect cannot be explained by local, disparity-specific motion adaptation, and the properties of scale and position invariance imply higher-level origins of neural adaptation. Nor is disparity essential for producing adaptation: when suspended on top of a stereoscopically defined, rotating globe, a context-disambiguated "globetrotter" was sufficient to bias the bistable walker's direction, as were full-body adaptors. In sum, these results imply that the neural signals supporting biomotion perception integrate information on the form, motion, and three-dimensional depth orientation of the moving human figure. Models of biomotion perception should incorporate mechanisms to disambiguate depth ambiguities in two-dimensional body representations.
Figures
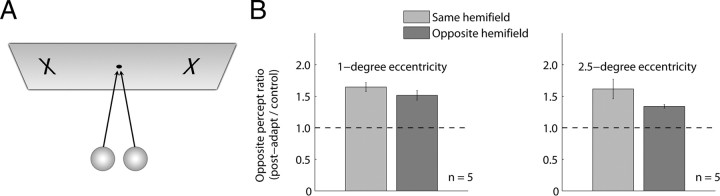
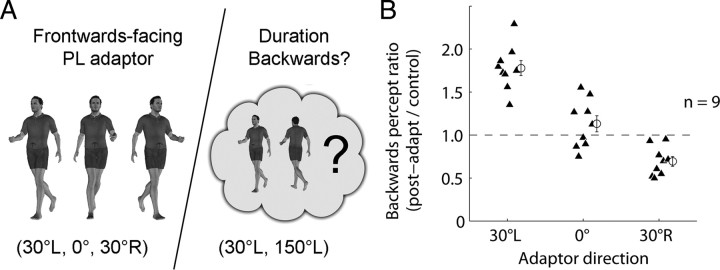
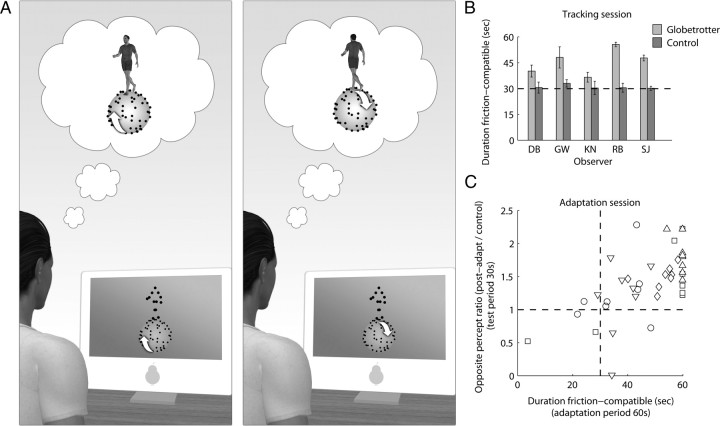
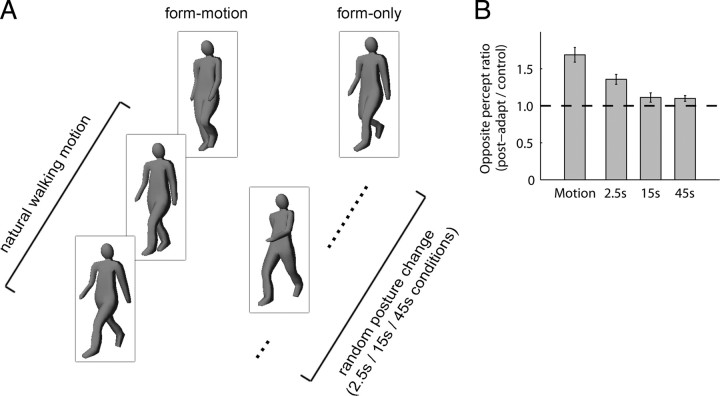
Similar articles
-
Stabilized structure from motion without disparity induces disparity adaptation.Curr Biol. 2004 Feb 3;14(3):247-51. doi: 10.1016/j.cub.2004.01.031. Curr Biol. 2004. PMID: 14761659
-
Perceptual adaptation to structure-from-motion depends on the size of adaptor and probe objects, but not on the similarity of their shapes.Atten Percept Psychophys. 2014 Feb;76(2):473-88. doi: 10.3758/s13414-013-0567-1. Atten Percept Psychophys. 2014. PMID: 24178065 Clinical Trial.
-
The stereoscopic (cyclopean) motion aftereffect is selective for spatial frequency and orientation of disparity modulation.Vision Res. 1999 Nov;39(22):3745-51. doi: 10.1016/s0042-6989(99)00087-5. Vision Res. 1999. PMID: 10746145
-
The aperture problem in egocentric motion.Trends Neurosci. 2004 Apr;27(4):174-7. doi: 10.1016/j.tins.2004.01.009. Trends Neurosci. 2004. PMID: 15108685 Review.
-
The motion aftereffect reloaded.Trends Cogn Sci. 2008 Dec;12(12):481-7. doi: 10.1016/j.tics.2008.09.002. Epub 2008 Oct 24. Trends Cogn Sci. 2008. PMID: 18951829 Free PMC article. Review.
Cited by
-
Individual differences in the perception of biological motion and fragmented figures are not correlated.Front Psychol. 2013 Oct 30;4:795. doi: 10.3389/fpsyg.2013.00795. eCollection 2013. Front Psychol. 2013. PMID: 24198799 Free PMC article.
-
Opposite influence of perceptual memory on initial and prolonged perception of sensory ambiguity.PLoS One. 2012;7(1):e30595. doi: 10.1371/journal.pone.0030595. Epub 2012 Jan 25. PLoS One. 2012. PMID: 22295095 Free PMC article.
-
Spatial selectivity in adaptation to gaze direction.Proc Biol Sci. 2022 Aug 10;289(1980):20221230. doi: 10.1098/rspb.2022.1230. Epub 2022 Aug 10. Proc Biol Sci. 2022. PMID: 35946160 Free PMC article.
-
Can biological motion research provide insight on how to reduce friendly fire incidents?Psychon Bull Rev. 2016 Oct;23(5):1429-1439. doi: 10.3758/s13423-016-1006-9. Psychon Bull Rev. 2016. PMID: 26850024 Review.
-
Biological movement and the encoding of its motion and orientation.Sci Rep. 2016 Feb 29;6:22393. doi: 10.1038/srep22393. Sci Rep. 2016. PMID: 26925870 Free PMC article.
References
-
- Barraclough NE, Xiao D, Oram MW, Perrett DI. The sensitivity of primate STS neurons to walking sequences and to the degree of articulation in static images. Prog Brain Res. 2006;154:135–148. - PubMed
-
- Battelli L, Cavanagh P, Thornton IM. Perception of biological motion in parietal patients. Neuropsychologia. 2003;41:1808–1816. - PubMed
-
- Blake R, Shiffrar M. Perception of human motion. Annu Rev Psychol. 2007;58:47–73. - PubMed
-
- Blakemore C, Julesz B. Stereoscopic depth aftereffect produced without monocular cues. Science. 1971;171:286–288. - PubMed
-
- Blakemore C, Sutton P. Size adaptation: a new aftereffect. Science. 1969;166:245–247. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources