

Structure of the HCMV UL16-MICB complex elucidates select binding of a viral immunoevasin to diverse NKG2D ligands
- PMID: 20090832
- PMCID: PMC2797645
- DOI: 10.1371/journal.ppat.1000723
Structure of the HCMV UL16-MICB complex elucidates select binding of a viral immunoevasin to diverse NKG2D ligands
Abstract
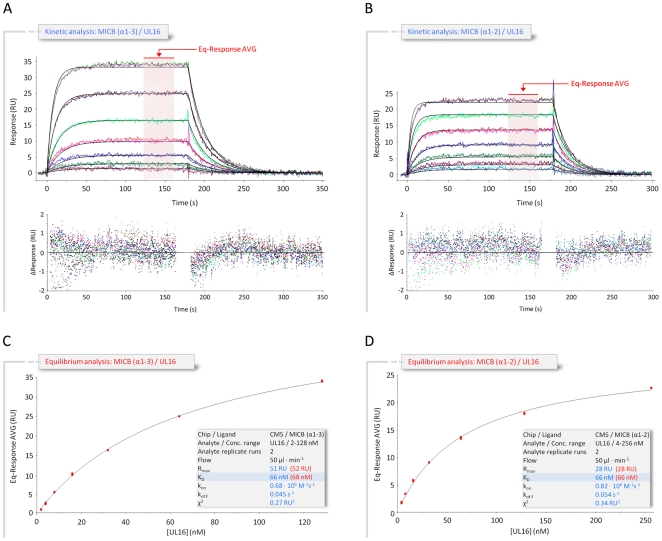
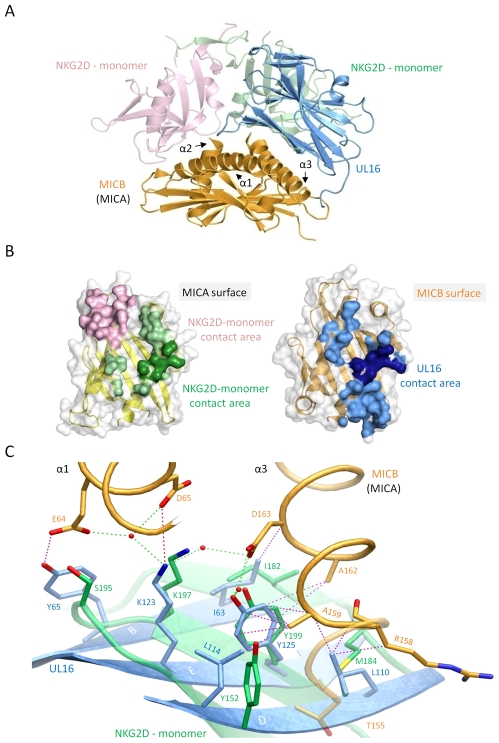
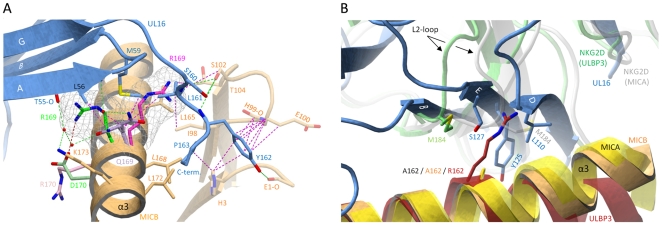
The activating immunoreceptor NKG2D promotes elimination of infected or malignant cells by cytotoxic lymphocytes through engagement of stress-induced MHC class I-related ligands. The human cytomegalovirus (HCMV)-encoded immunoevasin UL16 subverts NKG2D-mediated immune responses by retaining a select group of diverse NKG2D ligands inside the cell. We report here the crystal structure of UL16 in complex with the NKG2D ligand MICB at 1.8 A resolution, revealing the molecular basis for the promiscuous, but highly selective, binding of UL16 to unrelated NKG2D ligands. The immunoglobulin-like UL16 protein utilizes a three-stranded beta-sheet to engage the alpha-helical surface of the MHC class I-like MICB platform domain. Intriguingly, residues at the center of this beta-sheet mimic a central binding motif employed by the structurally unrelated C-type lectin-like NKG2D to facilitate engagement of diverse NKG2D ligands. Using surface plasmon resonance, we find that UL16 binds MICB, ULBP1, and ULBP2 with similar affinities that lie in the nanomolar range (12-66 nM). The ability of UL16 to bind its ligands depends critically on the presence of a glutamine (MICB) or closely related glutamate (ULBP1 and ULBP2) at position 169. An arginine residue at this position however, as found for example in MICA or ULBP3, would cause steric clashes with UL16 residues. The inability of UL16 to bind MICA and ULBP3 can therefore be attributed to single substitutions at key NKG2D ligand locations. This indicates that selective pressure exerted by viral immunoevasins such as UL16 contributed to the diversification of NKG2D ligands.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Effects of human cytomegalovirus infection on ligands for the activating NKG2D receptor of NK cells: up-regulation of UL16-binding protein (ULBP)1 and ULBP2 is counteracted by the viral UL16 protein.J Immunol. 2003 Jul 15;171(2):902-8. doi: 10.4049/jimmunol.171.2.902. J Immunol. 2003. PMID: 12847260
-
Human cytomegalovirus-encoded UL16 discriminates MIC molecules by their alpha2 domains.J Immunol. 2006 Sep 1;177(5):3143-9. doi: 10.4049/jimmunol.177.5.3143. J Immunol. 2006. PMID: 16920952
-
Human cytomegalovirus glycoprotein UL16 causes intracellular sequestration of NKG2D ligands, protecting against natural killer cell cytotoxicity.J Exp Med. 2003 Jun 2;197(11):1427-39. doi: 10.1084/jem.20022059. J Exp Med. 2003. PMID: 12782710 Free PMC article.
-
The UL16-binding proteins, a novel family of MHC class I-related ligands for NKG2D, activate natural killer cell functions.Immunol Rev. 2001 Jun;181:185-92. doi: 10.1034/j.1600-065x.2001.1810115.x. Immunol Rev. 2001. PMID: 11513139 Review.
-
Generation of soluble NKG2D ligands: proteolytic cleavage, exosome secretion and functional implications.Scand J Immunol. 2013 Aug;78(2):120-9. doi: 10.1111/sji.12072. Scand J Immunol. 2013. PMID: 23679194 Review.
Cited by
-
Allelic MHC class I chain related B (MICB) molecules affect the binding to the human cytomegalovirus (HCMV) unique long 16 (UL16) protein: implications for immune surveillance.J Microbiol. 2013 Apr;51(2):241-6. doi: 10.1007/s12275-013-2514-1. Epub 2013 Apr 27. J Microbiol. 2013. PMID: 23625227
-
Global Mapping of O-Glycosylation of Varicella Zoster Virus, Human Cytomegalovirus, and Epstein-Barr Virus.J Biol Chem. 2016 Jun 3;291(23):12014-28. doi: 10.1074/jbc.M116.721746. Epub 2016 Apr 15. J Biol Chem. 2016. PMID: 27129252 Free PMC article.
-
Structural basis for recognition of cellular and viral ligands by NK cell receptors.Front Immunol. 2014 Mar 26;5:123. doi: 10.3389/fimmu.2014.00123. eCollection 2014. Front Immunol. 2014. PMID: 24723923 Free PMC article. Review.
-
Recognition of Self and Viral Ligands by NK Cell Receptors.Immunol Rev. 2025 Jan;329(1):e13435. doi: 10.1111/imr.13435. Immunol Rev. 2025. PMID: 39748148 Free PMC article. Review.
-
Structural Models for Roseolovirus U20 And U21: Non-Classical MHC-I Like Proteins From HHV-6A, HHV-6B, and HHV-7.Front Immunol. 2022 Apr 4;13:864898. doi: 10.3389/fimmu.2022.864898. eCollection 2022. Front Immunol. 2022. PMID: 35444636 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
