

A proteomic approach for comprehensively screening substrates of protein kinases such as Rho-kinase
- PMID: 20090853
- PMCID: PMC2806833
- DOI: 10.1371/journal.pone.0008704
A proteomic approach for comprehensively screening substrates of protein kinases such as Rho-kinase
Abstract
Background: Protein kinases are major components of signal transduction pathways in multiple cellular processes. Kinases directly interact with and phosphorylate downstream substrates, thus modulating their functions. Despite the importance of identifying substrates in order to more fully understand the signaling network of respective kinases, efficient methods to search for substrates remain poorly explored.
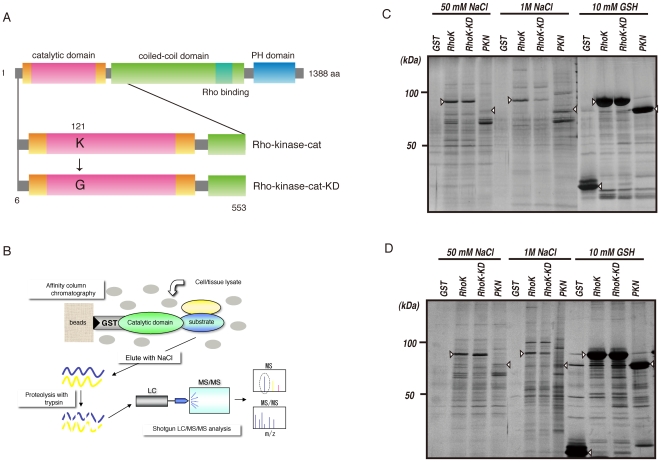
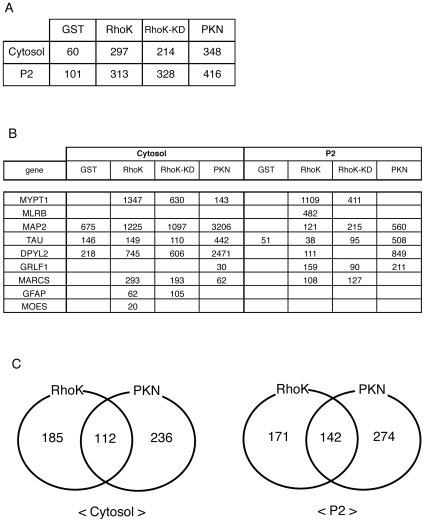
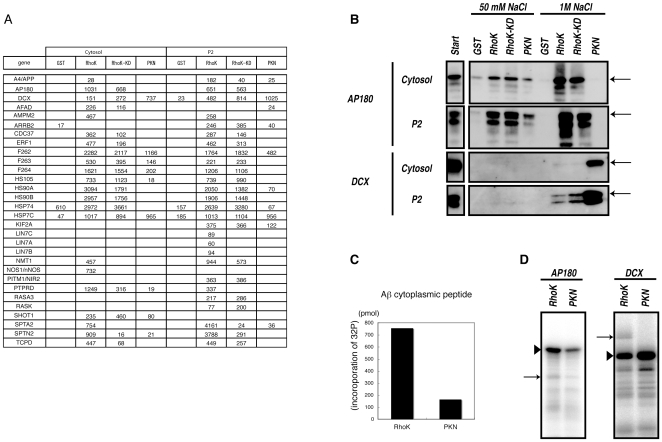
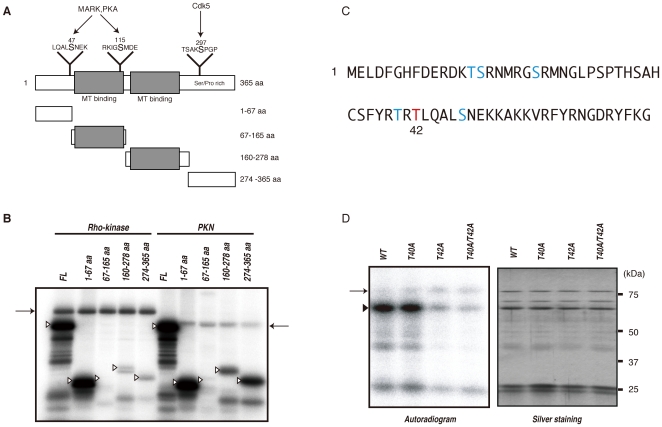
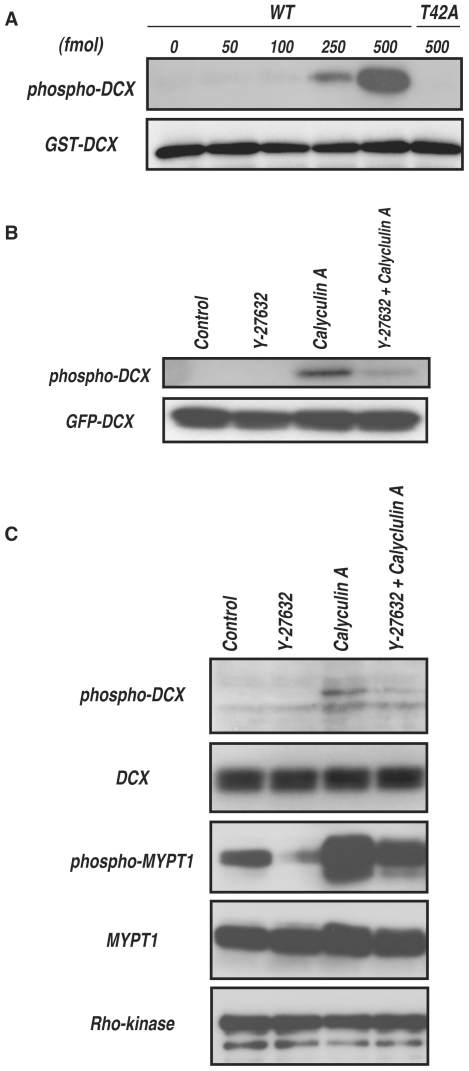
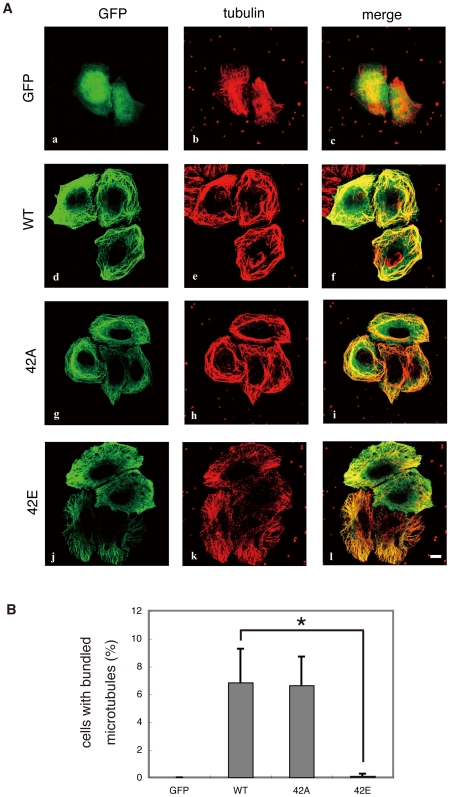
Methodology/principal findings: We combined mass spectrometry and affinity column chromatography of the catalytic domain of protein kinases to screen potential substrates. Using the active catalytic fragment of Rho-kinase/ROCK/ROK as the model bait, we obtained about 300 interacting proteins from the rat brain cytosol fraction, which included the proteins previously reported as Rho-kinase substrates. Several novel interacting proteins, including doublecortin, were phosphorylated by Rho-kinase both in vitro and in vivo.
Conclusions/significance: This method would enable identification of novel specific substrates for kinases such as Rho-kinase with high sensitivity.
Conflict of interest statement
Figures
References
-
- Ptacek J, Devgan G, Michaud G, Zhu H, Zhu X, et al. Global analysis of protein phosphorylation in yeast. Nature. 2005;438:679–684. - PubMed
-
- Schmelzle K, White FM. Phosphoproteomic approaches to elucidate cellular signaling networks. Curr Opin Biotechnol. 2006;17:406–414. - PubMed
-
- Tedford NC, Hall AB, Graham JR, Murphy CE, Gordon NF, et al. Quantitative analysis of cell signaling and drug action via mass spectrometry-based systems level phosphoproteomics. Proteomics. 2009;9:1469–1487. - PubMed
-
- Machida M, Kosako H, Shirakabe K, Kobayashi M, Ushiyama M, et al. Purification of phosphoproteins by immobilized metal affinity chromatography and its application to phosphoproteome analysis. Febs J. 2007;274:1576–1587. - PubMed
-
- Ubersax JA, Ferrell JE., Jr Mechanisms of specificity in protein phosphorylation. Nat Rev Mol Cell Biol. 2007;8:530–541. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
