

Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species
- PMID: 20093177
- PMCID: PMC2860730
- DOI: 10.1016/j.freeradbiomed.2010.01.015
Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species
Abstract
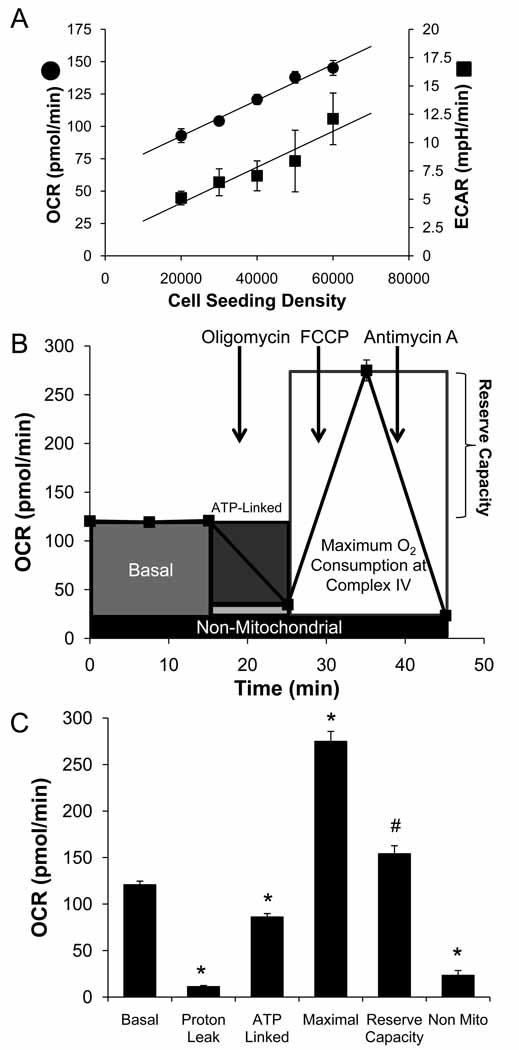
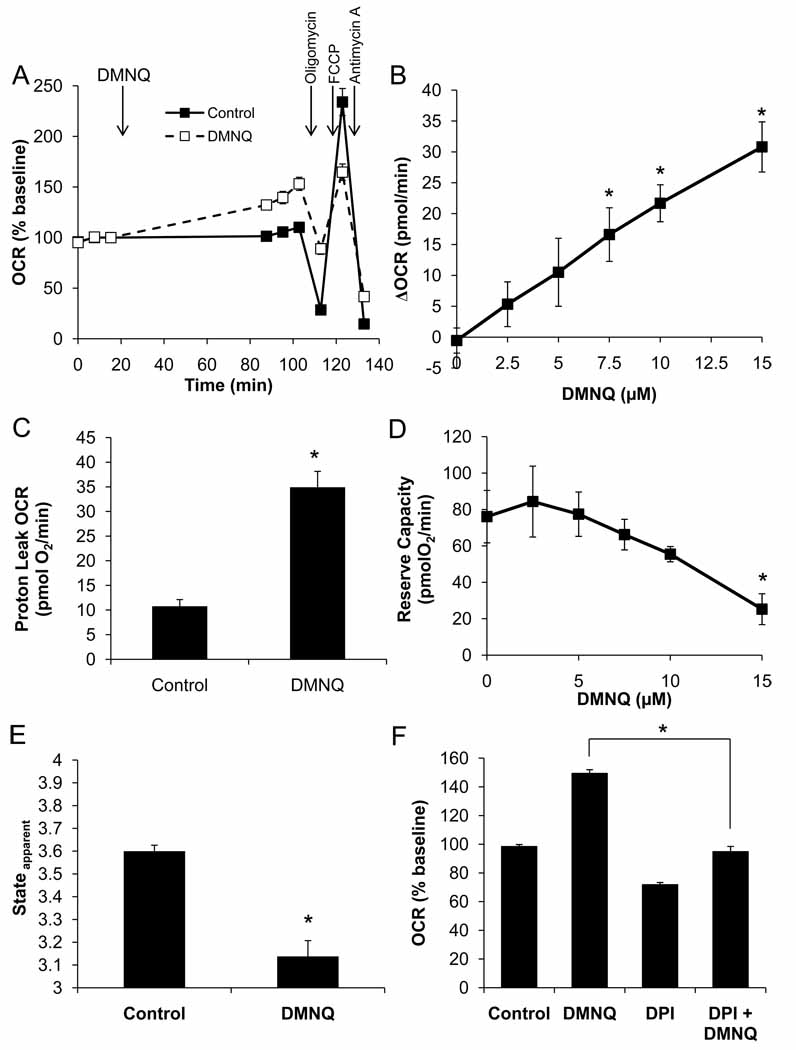
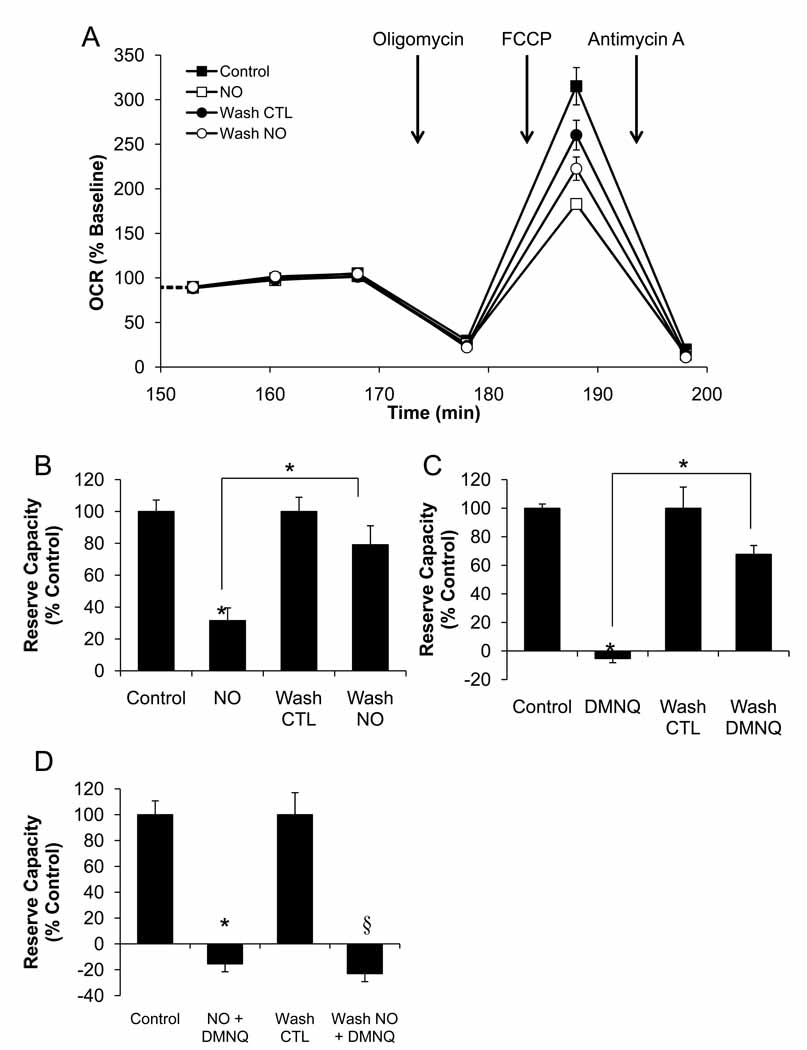
The endothelium is not considered to be a major energy-requiring organ, but nevertheless endothelial cells have an extensive mitochondrial network. This suggests that mitochondrial function may be important in response to stress and signaling in these cells. In this study, we used extracellular flux analysis to measure mitochondrial function in adherent bovine aortic endothelial cells (BAEC). Under basal conditions, BAEC use only approximately 35% of their maximal respiratory capacity. We calculate that this represents an intermediate respiratory state between States 3 and 4, which we define as State(apparent) equal to 3.64. Interestingly, the apparent respiratory control ratio (maximal mitochondrial oxygen consumption/non-ADP-linked respiration) in these cells is on the order of 23, which is substantially higher than that which is frequently obtained with isolated mitochondria. These results suggest that mitochondria in endothelial cells are highly coupled and possess a considerable bioenergetic reserve. Because endothelial cells are exposed to both reactive oxygen (ROS) and reactive nitrogen species in the course of vascular disease, we hypothesized that this reserve capacity is important in responding to oxidative stress. To test this, we exposed BAEC to NO or ROS alone or in combination. We found that exposure to nontoxic concentrations of NO or low levels of hydrogen peroxide generated from 2,3-dimethoxy-1,4-napthoquinone (DMNQ) had little impact on basal mitochondrial function but both treatments reversibly decreased mitochondrial reserve capacity. However, combined NO and DMNQ treatment resulted in an irreversible loss of reserve capacity and was associated with cell death. These data are consistent with a critical role for the mitochondrial reserve capacity in endothelial cells in responding to oxidative stress.
2010 Elsevier Inc. All rights reserved.
Figures






References
-
- Palmer RMJ, Ferrige AG, Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;327(6122):524–526. - PubMed
-
- Ramachandran A, et al. Activation of c-Jun N-Terminal Kinase and Apoptosis in Endothelial Cells Mediated by Endogenous Generation of Hydrogen Peroxide. Biological Chemistry. 2002;383(3):693. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
