

The intronic splicing code: multiple factors involved in ATM pseudoexon definition
- PMID: 20094034
- PMCID: PMC2829169
- DOI: 10.1038/emboj.2009.397
The intronic splicing code: multiple factors involved in ATM pseudoexon definition
Abstract
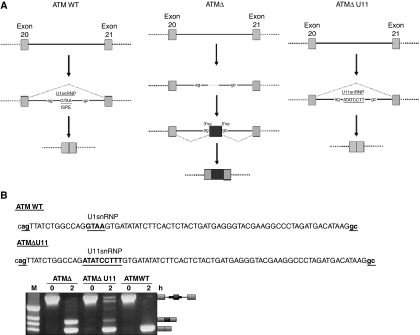
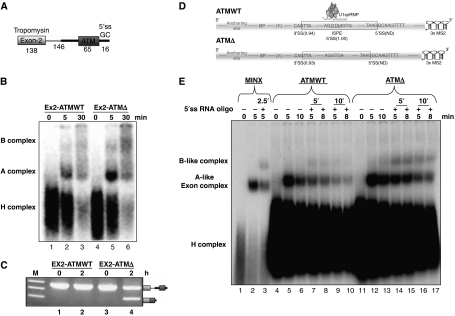
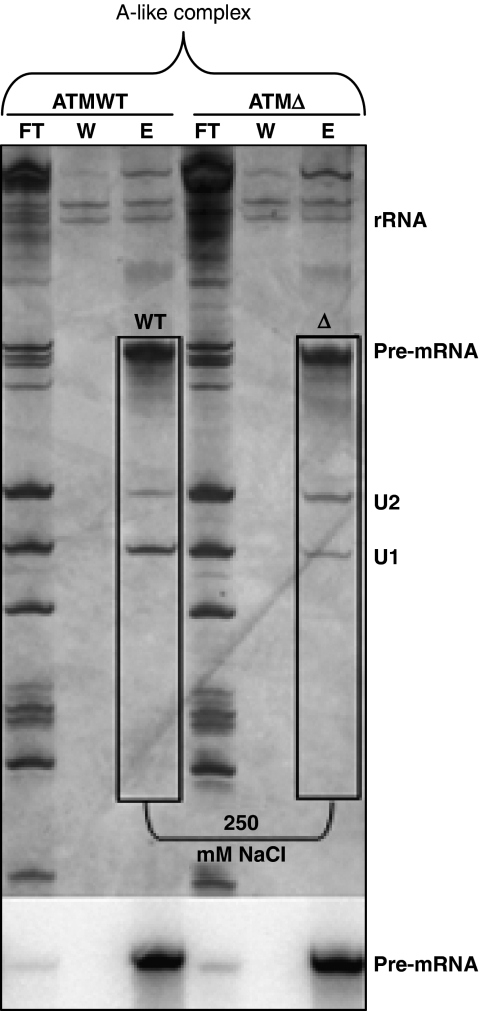
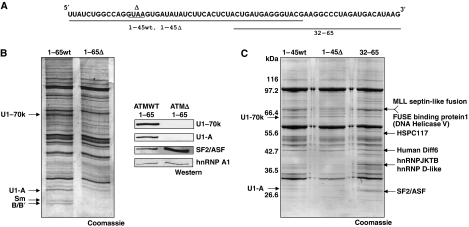
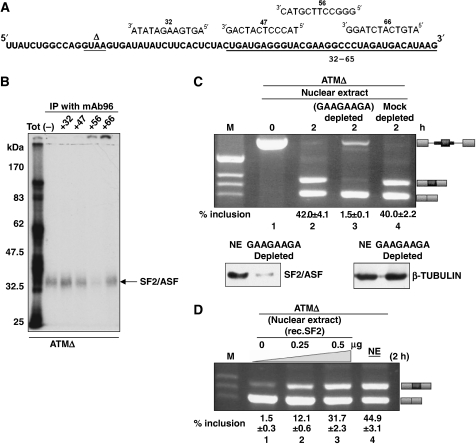
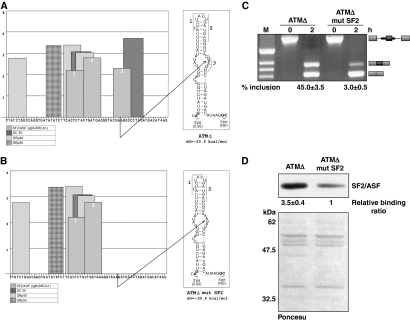
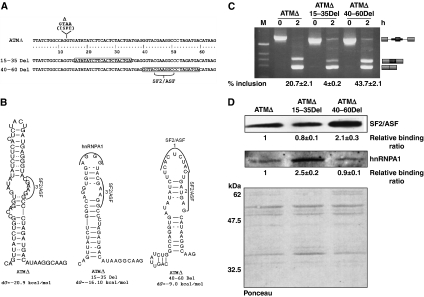
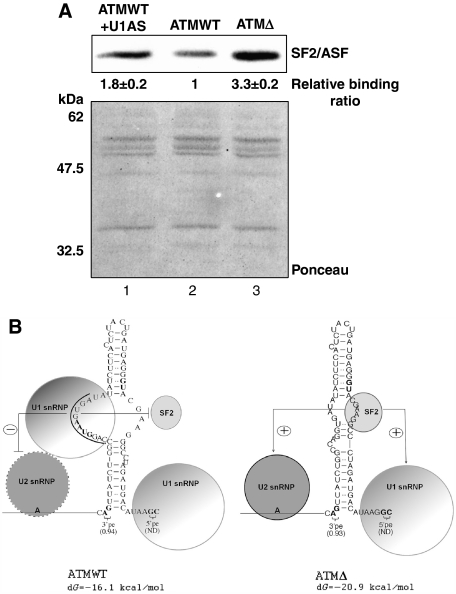
Abundance of pseudo splice sites in introns can potentially give rise to innumerable pseudoexons, outnumbering the real ones. Nonetheless, these are efficiently ignored by the splicing machinery, a process yet to be understood completely. Although numerous 5' splice site-like sequences functioning as splicing silencers have been found to be enriched in predicted human pseudoexons, the lack of active pseudoexons pose a fundamental challenge to how these U1snRNP-binding sites function in splicing inhibition. Here, we address this issue by focusing on a previously described pathological ATM pseudoexon whose inhibition is mediated by U1snRNP binding at intronic splicing processing element (ISPE), composed of a consensus donor splice site. Spliceosomal complex assembly demonstrates inefficient A complex formation when ISPE is intact, implying U1snRNP-mediated unproductive U2snRNP recruitment. Furthermore, interaction of SF2/ASF with its motif seems to be dependent on RNA structure and U1snRNP interaction. Our results suggest a complex combinatorial interplay of RNA structure and trans-acting factors in determining the splicing outcome and contribute to understanding the intronic splicing code for the ATM pseudoexon.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Functional studies on the ATM intronic splicing processing element.Nucleic Acids Res. 2005 Jul 19;33(13):4007-15. doi: 10.1093/nar/gki710. Print 2005. Nucleic Acids Res. 2005. PMID: 16030351 Free PMC article.
-
RNA structure is a key regulatory element in pathological ATM and CFTR pseudoexon inclusion events.Nucleic Acids Res. 2007;35(13):4369-83. doi: 10.1093/nar/gkm447. Epub 2007 Jun 18. Nucleic Acids Res. 2007. PMID: 17580311 Free PMC article.
-
Interactions of SR45, an SR-like protein, with spliceosomal proteins and an intronic sequence: insights into regulated splicing.Plant J. 2012 Sep;71(6):936-47. doi: 10.1111/j.1365-313X.2012.05042.x. Epub 2012 Jun 28. Plant J. 2012. PMID: 22563826
-
Phosphorylation-dependent control of the pre-mRNA splicing machinery.Prog Mol Subcell Biol. 2003;31:89-126. doi: 10.1007/978-3-662-09728-1_4. Prog Mol Subcell Biol. 2003. PMID: 12494764 Review. No abstract available.
-
Branch site recognition by the spliceosome.RNA. 2024 Oct 16;30(11):1397-1407. doi: 10.1261/rna.080198.124. RNA. 2024. PMID: 39187383 Free PMC article. Review.
Cited by
-
Alternative splicing of U2AF1 reveals a shared repression mechanism for duplicated exons.Nucleic Acids Res. 2017 Jan 9;45(1):417-434. doi: 10.1093/nar/gkw733. Epub 2016 Aug 26. Nucleic Acids Res. 2017. PMID: 27566151 Free PMC article.
-
Upon the tightrope in prostate cancer: two acrobats on the same tightrope to cross the finishline.Mol Cell Biochem. 2012 May;364(1-2):53-7. doi: 10.1007/s11010-011-1204-8. Epub 2011 Dec 27. Mol Cell Biochem. 2012. PMID: 22200977 Review.
-
A novel splicing silencer generated by DMD exon 45 deletion junction could explain upstream exon 44 skipping that modifies dystrophinopathy.J Hum Genet. 2014 Aug;59(8):423-9. doi: 10.1038/jhg.2014.36. Epub 2014 May 29. J Hum Genet. 2014. PMID: 24871807
-
Role of Splicing Regulatory Elements and In Silico Tools Usage in the Identification of Deep Intronic Splicing Variants in Hereditary Breast/Ovarian Cancer Genes.Cancers (Basel). 2021 Jul 3;13(13):3341. doi: 10.3390/cancers13133341. Cancers (Basel). 2021. PMID: 34283047 Free PMC article.
-
A Deep Intronic Variant Activates a Pseudoexon in the MTM1 Gene in a Family with X-Linked Myotubular Myopathy.Mol Syndromol. 2020 Dec;11(5-6):264-270. doi: 10.1159/000510286. Epub 2020 Sep 16. Mol Syndromol. 2020. PMID: 33505229 Free PMC article.
References
-
- Akker SA, Misra S, Aslam S, Morgan EL, Smith PJ, Khoo B, Chew SL (2007) Pre-spliceosomal binding of U1 small nuclear ribonucleoprotein (RNP) and heterogenous nuclear RNP E1 is associated with suppression of a growth hormone receptor pseudoexon. Mol Endocrinol 21: 2529–2540 - PubMed
-
- Amara SG, Jonas V, Rosenfeld MG, Ong ES, Evans RM (1982) Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature 298: 240–244 - PubMed
-
- Baralle M, Skoko N, Knezevich A, De Conti L, Motti D, Bhuvanagiri M, Baralle D, Buratti E, Baralle FE (2006) NF1 mRNA biogenesis: effect of the genomic milieu in splicing regulation of the NF1 exon 37 region. FEBS Lett 580: 4449–4456 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
