

A study of cytochrome bo3 in a tethered bilayer lipid membrane
- PMID: 20096262
- PMCID: PMC3827738
- DOI: 10.1016/j.bbabio.2010.01.012
A study of cytochrome bo3 in a tethered bilayer lipid membrane
Abstract
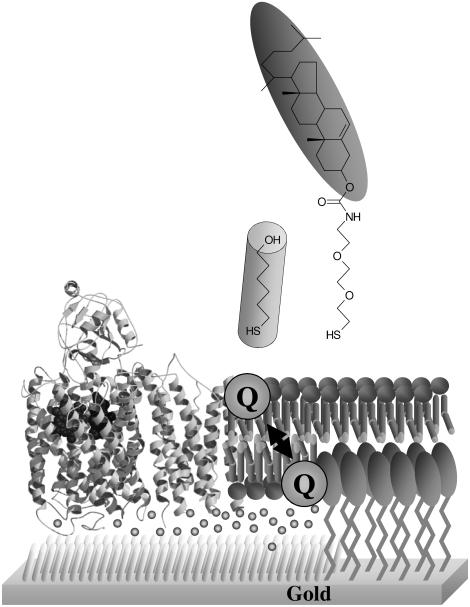
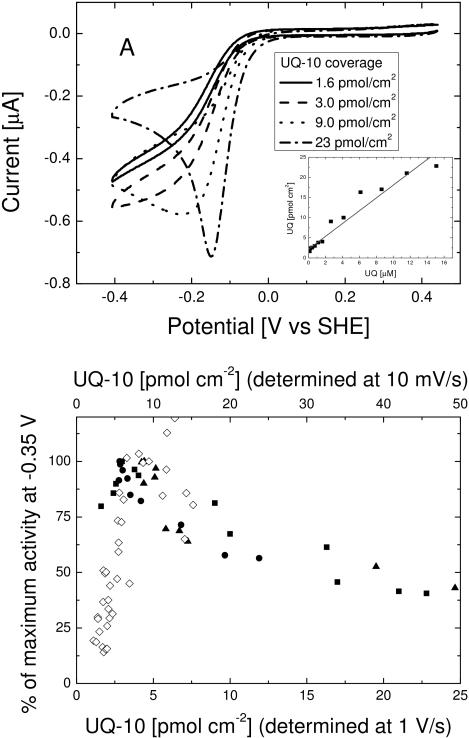
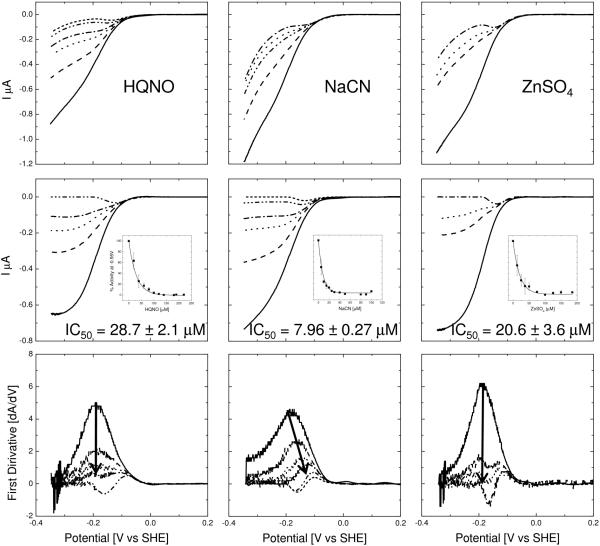
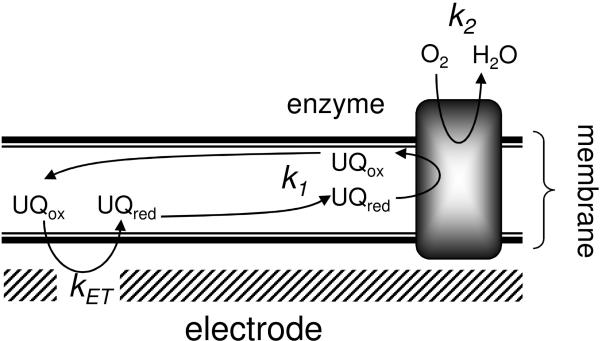
An assay has been developed in which the activity of an ubiquinol oxidase from Escherichia coli, cytochrome bo(3) (cbo(3)), is determined as a function of the hydrophobic substrate ubiquinol-10 (UQ-10) in tethered bilayer lipid membranes (tBLMs). UQ-10 was added in situ, while the enzyme activity and the UQ-10 concentration in the membrane have been determined by cyclic voltammetry. Cbo(3) is inhibited by UQ-10 at concentrations above 5-10 pmol/cm(2), while product inhibition is absent. Cyclic voltammetry has also been used to characterise the effects of three inhibitors; cyanide, inhibiting oxygen reduction; 2-n-Heptyl-4-hydroxyquinoline N-oxide (HQNO), inhibiting the quinone oxidation and Zn(II), thought to block the proton channels required for oxygen reduction and proton pumping activity. The electrochemical behaviour of cbo(3) inhibited with HQNO and Zn(II) is almost identical, suggesting that Zn(II) ions inhibit the enzyme reduction by quinol, rather than oxygen reduction. This suggests that at Zn(II) concentration below 50µM the proton release of cbo(3) is inhibited, but not the proton uptake required to reduce oxygen to water.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
The quinone-binding sites of the cytochrome bo3 ubiquinol oxidase from Escherichia coli.Biochim Biophys Acta. 2010 Dec;1797(12):1924-32. doi: 10.1016/j.bbabio.2010.04.011. Epub 2010 Apr 20. Biochim Biophys Acta. 2010. PMID: 20416270 Free PMC article.
-
Uncompetitive substrate inhibition and noncompetitive inhibition by 5-n-undecyl-6-hydroxy-4,7-dioxobenzothiazole (UHDBT) and 2-n-nonyl-4-hydroxyquinoline-N-oxide (NQNO) is observed for the cytochrome bo3 complex: implications for a Q(H2)-loop proton translocation mechanism.Biochemistry. 1997 Jan 28;36(4):894-902. doi: 10.1021/bi961723r. Biochemistry. 1997. PMID: 9020789
-
Characterization of cytochrome bo3 activity in a native-like surface-tethered membrane.Biochem J. 2009 Jan 15;417(2):555-60. doi: 10.1042/BJ20081345. Biochem J. 2009. PMID: 18821852 Free PMC article.
-
Redox enzymes in tethered membranes.J Am Chem Soc. 2006 Feb 8;128(5):1711-6. doi: 10.1021/ja056972u. J Am Chem Soc. 2006. PMID: 16448146 Free PMC article.
-
Oxygen as Acceptor.EcoSal Plus. 2015;6(2):10.1128/ecosalplus.ESP-0012-2015. doi: 10.1128/ecosalplus.ESP-0012-2015. EcoSal Plus. 2015. PMID: 26734697 Free PMC article. Review.
Cited by
-
Evidence for distinct electron transfer processes in terminal oxidases from different origin by means of protein film voltammetry.J Am Chem Soc. 2014 Aug 6;136(31):10854-7. doi: 10.1021/ja505126v. Epub 2014 Jul 25. J Am Chem Soc. 2014. PMID: 25054669 Free PMC article.
-
Cryo-EM structures of Escherichia coli cytochrome bo3 reveal bound phospholipids and ubiquinone-8 in a dynamic substrate binding site.Proc Natl Acad Sci U S A. 2021 Aug 24;118(34):e2106750118. doi: 10.1073/pnas.2106750118. Proc Natl Acad Sci U S A. 2021. PMID: 34417297 Free PMC article.
-
The respiratory chain of Klebsiella aerogenes in urine-like conditions: critical roles of NDH-2 and bd-terminal oxidases.Front Microbiol. 2024 Nov 6;15:1479714. doi: 10.3389/fmicb.2024.1479714. eCollection 2024. Front Microbiol. 2024. PMID: 39568993 Free PMC article.
-
Direct Electrochemistry of Cytochrome bo Oxidase at a series of Gold Nanoparticles-Modified Electrodes.Electrochem commun. 2013 Jan 1;26:105-108. doi: 10.1016/j.elecom.2012.10.024. Epub 2012 Oct 23. Electrochem commun. 2013. PMID: 23335854 Free PMC article.
-
Solid-supported polymer-lipid hybrid membrane for bioelectrochemistry of a membrane redox enzyme.RSC Appl Interfaces. 2025 Feb 11;2(3):665-672. doi: 10.1039/d4lf00362d. eCollection 2025 May 14. RSC Appl Interfaces. 2025. PMID: 39980607 Free PMC article.
References
-
- Anraku Y. Bacterial Electron-Transport Chains. Annu. Rev. Biochem. 1988;57:101–132. - PubMed
-
- Soballe B, Poole RK. Microbial ubiquinones: multiple roles in respiration, gene regulation and oxidative stress management. Microbiology-(UK) 1999;145:1817–1830. - PubMed
-
- Kita K, Konishi K, Anraku Y. Terminal oxidases of Escherichia coli aerobic respiratory chain .1. Purification and properties of cytochrome b562o complex from cells in the early exponential phase of aerobic growth. J. Biol. Chem. 1984;259:3368–3374. - PubMed
-
- Kita K, Konishi K, Anraku Y. Terminal oxidases of Escherichia coli aerobic respiratory chain .2. Purification and properties of cytochrome b558d complex from cells grown with limited oxygen and evidence of branched electron-carrying systems. J. Biol. Chem. 1984;259:3375–3381. - PubMed
-
- Abramson J, Riistama S, Larsson G, Jasaitis A, Svensson-Ek M, Laakkonen L, Puustinen A, Iwata S, Wikstrom M. The structure of the ubiquinol oxidase from Escherichia coli and its ubiquinone binding site. Nat. Struct. Biol. 2000;7:910–917. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
