

Review
doi: 10.1016/j.dnarep.2009.12.009.
Epub 2010 Jan 22.
Recruitment to stalled replication forks of the PriA DNA helicase and replisome-loading activities is essential for survival
Affiliations
- PMID: 20097140
- PMCID: PMC2827650
- DOI: 10.1016/j.dnarep.2009.12.009
Item in Clipboard
Review
Recruitment to stalled replication forks of the PriA DNA helicase and replisome-loading activities is essential for survival
DNA Repair (Amst).
.
Abstract
PriA, a 3'-->5' superfamily 2 DNA helicase, acts to remodel stalled replication forks and as a specificity factor for origin-independent assembly of a new replisome at the stalled fork. The ability of PriA to initiate replication at stalled forked structures ensures complete genome replication and helps to protect the cell from illegitimate recombination events. This review focuses on the activities of PriA and its role in replication fork assembly and maintaining genomic integrity.
(c) 2010 Elsevier B.V. All rights reserved.
Figures
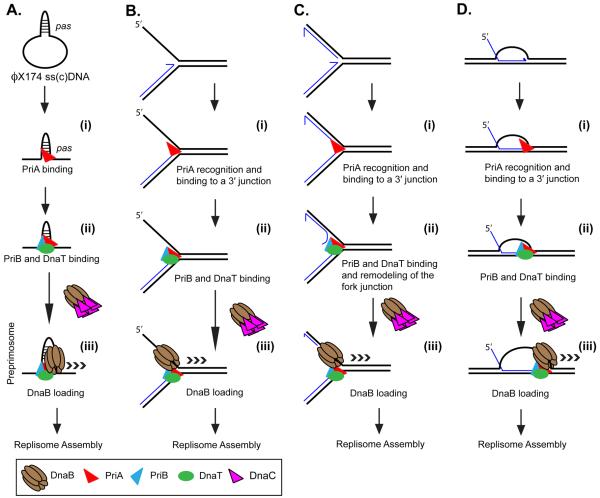
PriA replisome loading. PriA can load a replisome to various DNA structures. (A) A primosome assembly site on ϕX174 viral DNA. (B) A stalled replication fork where there is a gap in the nascent lagging-strand. Such a structure can form when a leading-strand template block terminates all DNA synthesis. (C) A stalled replication fork where both template strands have been completely copied up to the point of template damage. Similar to (B) above, such a structure may occur when the replisome encounters a leading-strand blockage. Replisome loading on such a structure requires the unwinding of a portion of the nascent lagging-strand DNA to generate a single-stranded region for DnaB binding. (D) A recombinant joint molecule, represented here as a D loop. These structures necessarily occur for restart after the encounter of a template double-strand break and may also be generated under other situations as well. Although the DNA structures are different, the steps in loading the replisome are identical. (i) PriA recognizes the 3′ OH end of the nascent leading-strand DNA at the three-strand junction and (ii) recruits other accessory factors that may be important in fork remodeling (for example, in panel C). Recognition of the pas probably occurs through interaction of PriA with the psuedo-three stand junction formed by the hairpin in the DNA and the adjacent single-stranded DNA. (iii) An interaction between DnaT and DnaC, the helicase loader, allows loading of the replicative helicase, DnaB, forming the preprimosome (composed of PriA, PriB, DnaT, and DnaB). Once loaded, DnaB recruits DnaG via a protein-protein interaction. Synthesis of a primer then attracts the DNA polymerase III holoenzyme and a protein-protein interaction between the τ subunit of the holoenzyme and DnaB cements formation of the replisome. Nascent template 3′ OH ends are denoted by half-arrows.
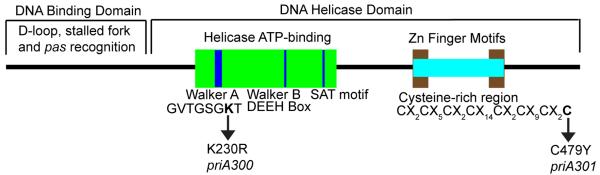
Amino acid motifs of E. coli PriA. PriA is divided into two major domains, a DNA-binding domain and a DNA helicase domain. The DNA helicase domain consists of an ATP-binding domain composed of a Walker A box, Walker B box, and SAT motif, and a cysteine-rich region that contains two Zn-finger motifs. Amino acid substitutions in the Walker A box, K230R, yielding a helicase-dead mutant protein, and in the metal binding domain, C479Y, representing the priA300 and priA301 mutations, are shown.
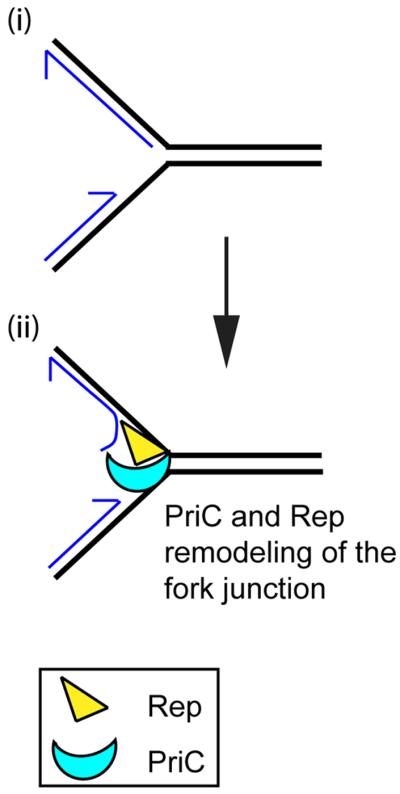
Remodeling of stalled replication forks during replisome loading by the PriC pathway. The preferred DNA structure (i) for the PriC pathway is likely formed when the replisome encounters a leading-strand template blockage and the leading- and lagging-strand polymerases of the replicase uncouple. This results in a gap in the nascent leading strand at the stalled fork with the 5′ of the nascent lagging strand being either right at or close to the fork junction. Displacement of the nascent lagging-strand to allow DnaB loading to the lagging-strand template is most likely accomplished by PriC directing Rep to the proper DNA strand (ii).
References
-
- Allen GC, Jr., Kornberg A. Assembly of the primosome of DNA replication in Escherichia coli. J Biol Chem. 1993;268:19204–9. - PubMed
-
- Arber W. Host-controlled variation. In: Hershey AD, editor. The Bacteriophage Lambda. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 1971. pp. 83–96.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
