

C-type lectin Langerin is a beta-glucan receptor on human Langerhans cells that recognizes opportunistic and pathogenic fungi
- PMID: 20097424
- PMCID: PMC2837148
- DOI: 10.1016/j.molimm.2009.12.016
C-type lectin Langerin is a beta-glucan receptor on human Langerhans cells that recognizes opportunistic and pathogenic fungi
Abstract
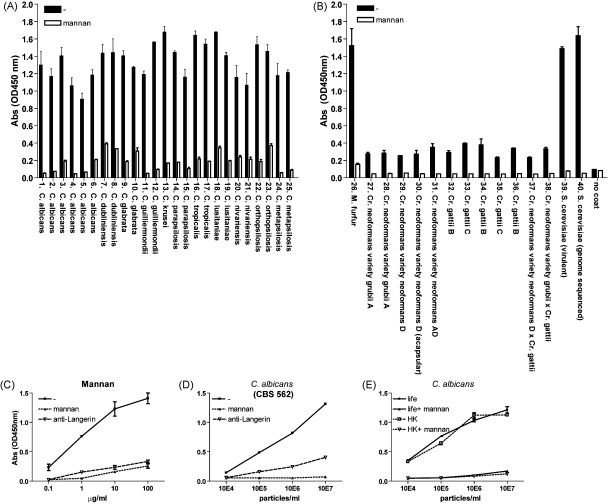
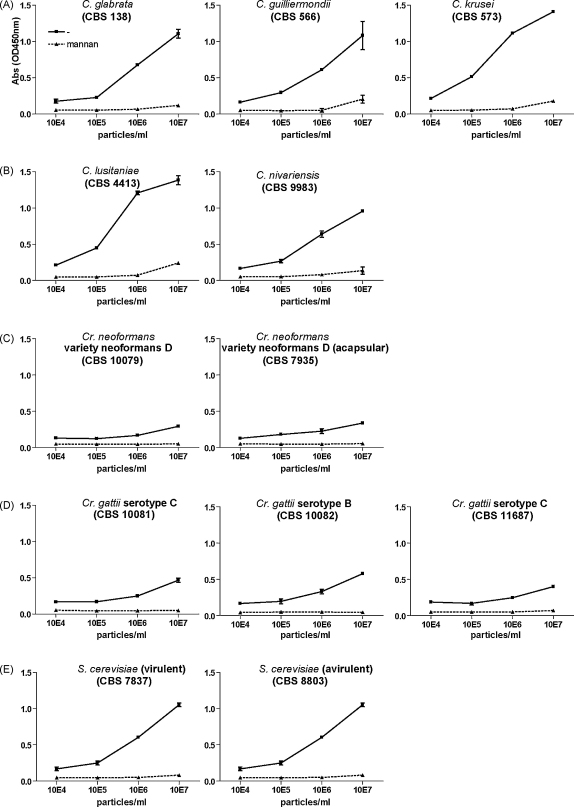
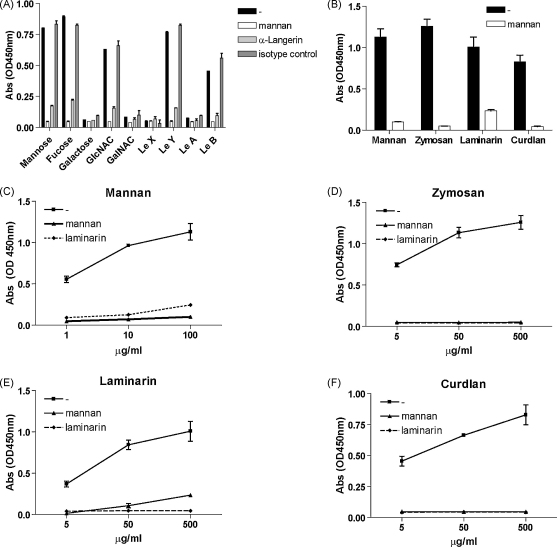
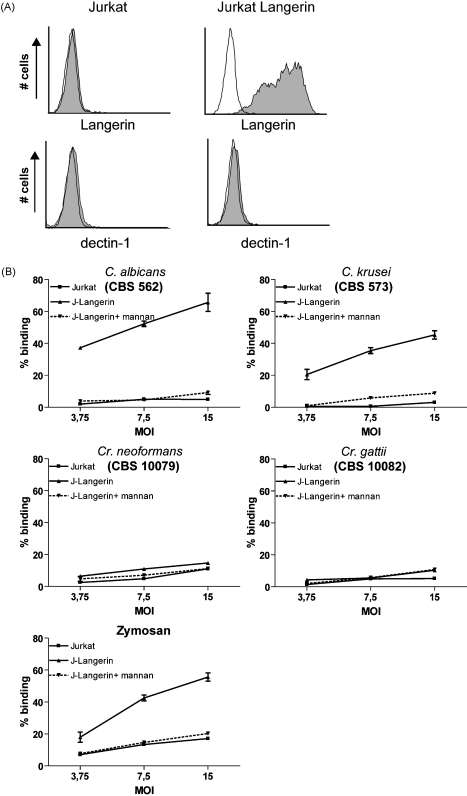
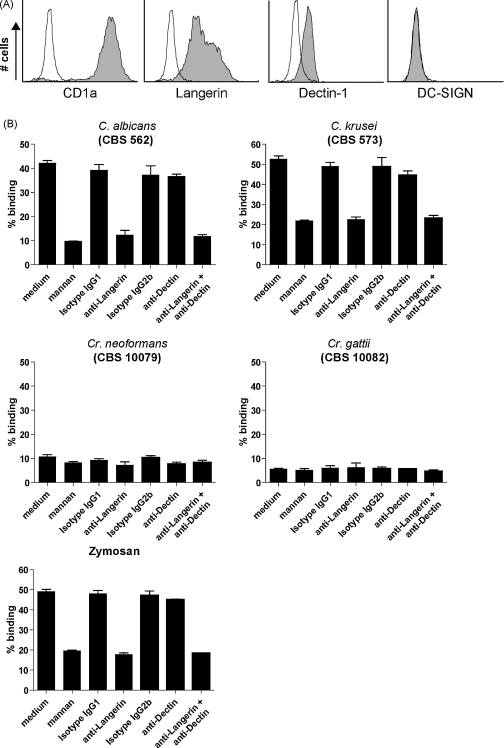
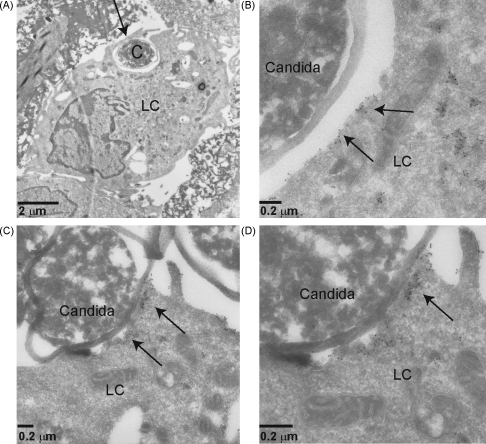
Langerhans cells (LCs) lining the stratified epithelia and mucosal tissues are the first antigen presenting cells to encounter invading pathogens, such as viruses, bacteria and fungi. Fungal infections form a health threat especially in immuno-compromised individuals. LCs express C-type lectin Langerin that has specificity for mannose, fucose and GlcNAc structures. Little is known about the role of human Langerin in fungal infections. Our data show that Langerin interacts with both mannan and beta-glucan structures, common cell-wall carbohydrate structures of fungi. We have screened a large panel of fungi for recognition by human Langerin and, strikingly, we observed strong binding of Langerin to a variety of Candida and Saccharomyces species and Malassezia furfur, but very weak binding was observed to Cryptococcus gattii and Cryptococcus neoformans. Notably, Langerin is the primary fungal receptor on LCs, since the interaction of LCs with the different fungi was blocked by antibodies against Langerin. Langerin recognizes both mannose and beta-glucans present on fungal cell walls and our data demonstrate that Langerin is the major fungal pathogen receptor on human LCs that recognizes pathogenic and commensal fungi. Together these data may provide more insight in the role of LCs in fungal infections.
Copyright 2009 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Dual specificity of Langerin to sulfated and mannosylated glycans via a single C-type carbohydrate recognition domain.J Biol Chem. 2010 Feb 26;285(9):6390-400. doi: 10.1074/jbc.M109.041863. Epub 2009 Dec 21. J Biol Chem. 2010. PMID: 20026605 Free PMC article.
-
Langerhans Cell-Dendritic Cell Cross-Talk via Langerin and Hyaluronic Acid Mediates Antigen Transfer and Cross-Presentation of HIV-1.J Immunol. 2015 Aug 15;195(4):1763-73. doi: 10.4049/jimmunol.1402356. Epub 2015 Jul 13. J Immunol. 2015. PMID: 26170391
-
Receptor usage dictates HIV-1 restriction by human TRIM5α in dendritic cell subsets.Nature. 2016 Dec 15;540(7633):448-452. doi: 10.1038/nature20567. Epub 2016 Dec 7. Nature. 2016. PMID: 27919079
-
Langerin functions as an antiviral receptor on Langerhans cells.Immunol Cell Biol. 2010 May-Jun;88(4):410-5. doi: 10.1038/icb.2010.32. Epub 2010 Mar 23. Immunol Cell Biol. 2010. PMID: 20309013 Review.
-
Langerin/CD207 sheds light on formation of birbeck granules and their possible function in Langerhans cells.Immunol Res. 2003;28(2):93-107. doi: 10.1385/IR:28:2:93. Immunol Res. 2003. PMID: 14610287 Review.
Cited by
-
Vulvovaginal Candidosis (Excluding Mucocutaneous Candidosis): Guideline of the German (DGGG), Austrian (OEGGG) and Swiss (SGGG) Society of Gynecology and Obstetrics (S2k-Level, AWMF Registry Number 015/072, September 2020).Geburtshilfe Frauenheilkd. 2021 Apr;81(4):398-421. doi: 10.1055/a-1345-8793. Epub 2021 Apr 14. Geburtshilfe Frauenheilkd. 2021. PMID: 33867561 Free PMC article.
-
Vulvovaginal Candidosis (excluding chronic mucocutaneous candidosis). Guideline of the German Society of Gynecology and Obstetrics (AWMF Registry No. 015/072, S2k Level, December 2013).Geburtshilfe Frauenheilkd. 2015 Apr;75(4):342-354. doi: 10.1055/s-0035-1545741. Geburtshilfe Frauenheilkd. 2015. PMID: 27065484 Free PMC article. No abstract available.
-
Common polymorphisms in human langerin change specificity for glycan ligands.J Biol Chem. 2013 Dec 27;288(52):36762-71. doi: 10.1074/jbc.M113.528000. Epub 2013 Nov 11. J Biol Chem. 2013. PMID: 24217250 Free PMC article.
-
Beta-1,3 Oligoglucans Specifically Bind to Immune Receptor CD28 and May Enhance T Cell Activation.Int J Mol Sci. 2021 Mar 18;22(6):3124. doi: 10.3390/ijms22063124. Int J Mol Sci. 2021. PMID: 33803858 Free PMC article.
-
C-Type Lectin Receptors in Host Defense Against Bacterial Pathogens.Front Cell Infect Microbiol. 2020 Jul 7;10:309. doi: 10.3389/fcimb.2020.00309. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32733813 Free PMC article. Review.
References
-
- Almirante B., Rodriguez D., Park B.J., Cuenca-Estrella M., Planes A.M., Almela M., Mensa J., Sanchez F., Ayats J., Gimenez M., Saballs P., Fridkin S.K., Morgan J., Rodriguez-Tudela J.L., Warnock D.W., Pahissa A. Epidemiology and predictors of mortality in cases of Candida bloodstream infection: results from population-based surveillance, barcelona, Spain, from 2002 to 2003. J. Clin. Microbiol. 2005;43:1829–1835. - PMC - PubMed
-
- Arendrup M.C., Fuursted K., Gahrn-Hansen B., Schonheyder H.C., Knudsen J.D., Jensen I.M., Bruun B., Christensen J.J., Johansen H.K. Semi-national surveillance of fungaemia in Denmark 2004–2006: increasing incidence of fungaemia and numbers of isolates with reduced azole susceptibility. Clin. Microbiol. Infect. 2008;14:487–494. - PubMed
-
- Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
-
- Batra R., Boekhout T., Gueho E., Cabanes F.J., Dawson T.L., Jr., Gupta A.K. Malassezia Baillon, emerging clinical yeasts. FEMS Yeast Res. 2005;5:1101–1113. - PubMed
-
- Bovers M., Hagen F., Kuramae E.E., Diaz M.R., Spanjaard L., Dromer F., Hoogveld H.L., Boekhout T. Unique hybrids between the fungal pathogens Cryptococcus neoformans and Cryptococcus gattii. FEMS Yeast Res. 2006;6:599–607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
