

Comparative transcriptome analysis reveals novel roles of the Ras and cyclic AMP signaling pathways in environmental stress response and antifungal drug sensitivity in Cryptococcus neoformans
- PMID: 20097740
- PMCID: PMC2837985
- DOI: 10.1128/EC.00309-09
Comparative transcriptome analysis reveals novel roles of the Ras and cyclic AMP signaling pathways in environmental stress response and antifungal drug sensitivity in Cryptococcus neoformans
Abstract
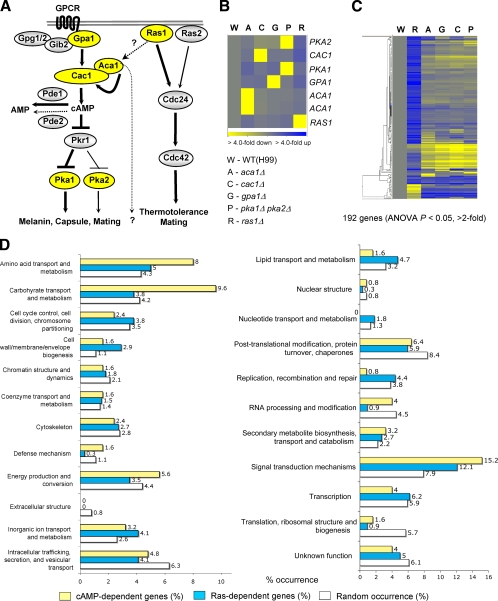
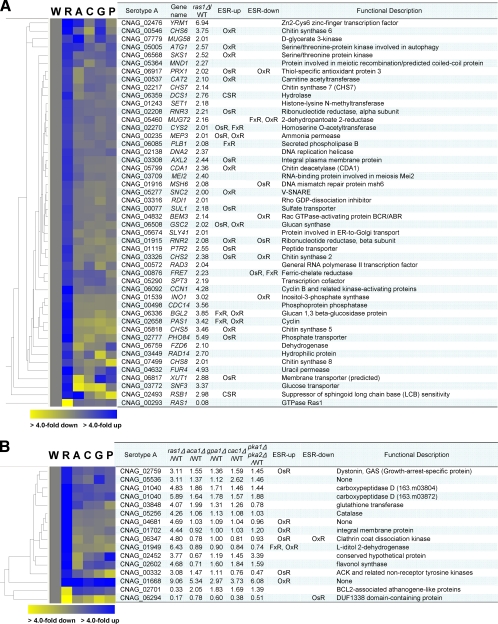
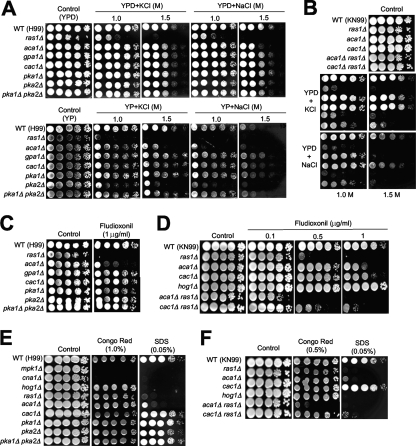
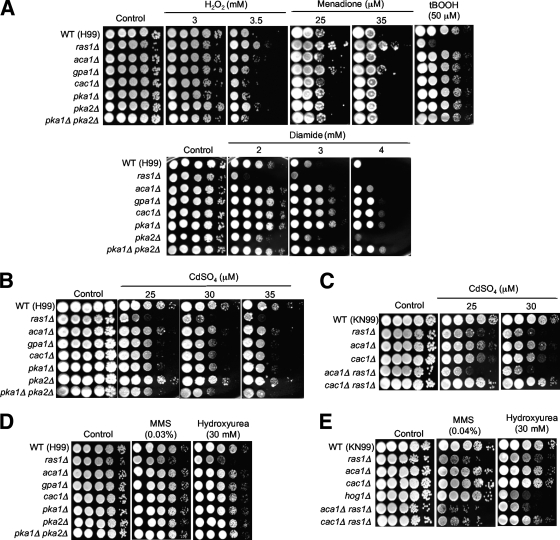
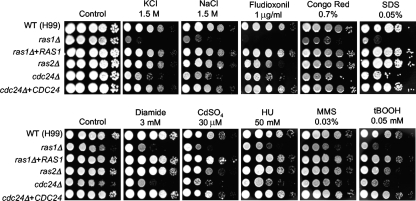
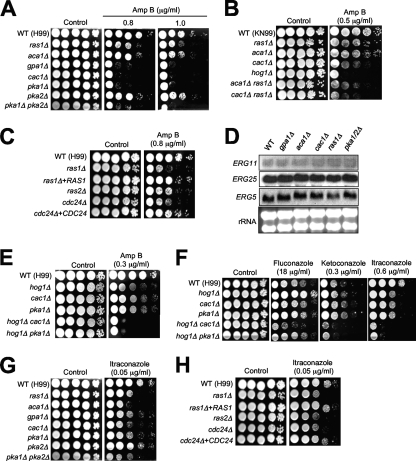
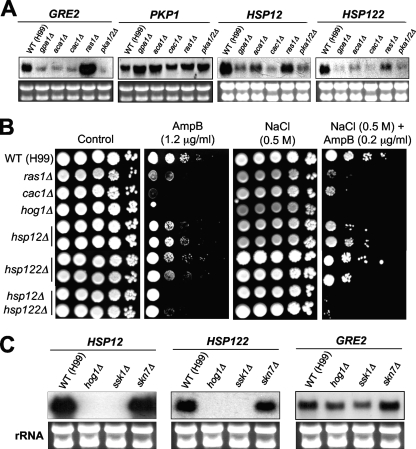
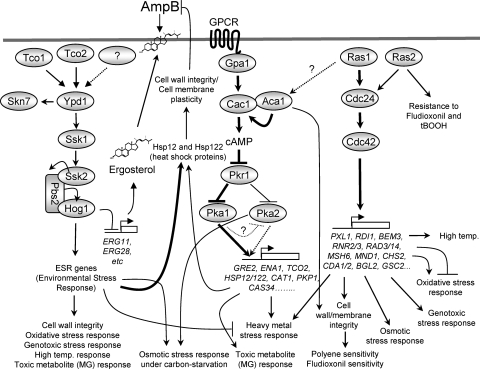
The cyclic AMP (cAMP) pathway plays a central role in the growth, differentiation, and virulence of pathogenic fungi, including Cryptococcus neoformans. Three upstream signaling regulators of adenylyl cyclase (Cac1), Ras, Aca1, and Gpa1, have been demonstrated to control the cAMP pathway in C. neoformans, but their functional relationship remains elusive. We performed a genome-wide transcriptome analysis with a DNA microarray using the ras1Delta, gpa1Delta, cac1Delta, aca1Delta, and pka1Delta pka2Delta mutants. The aca1Delta, gpa1Delta, cac1Delta, and pka1Delta pka2Delta mutants displayed similar transcriptome patterns, whereas the ras1Delta mutant exhibited transcriptome patterns distinct from those of the wild type and the cAMP mutants. Interestingly, a number of environmental stress response genes are modulated differentially in the ras1Delta and cAMP mutants. In fact, the Ras signaling pathway was found to be involved in osmotic and genotoxic stress responses and the maintenance of cell wall integrity via the Cdc24-dependent signaling pathway. Notably, the Ras and cAMP mutants exhibited hypersensitivity to a polyene drug, amphotericin B, without showing effects on ergosterol biosynthesis, which suggested a novel method of antifungal combination therapy. Among the cAMP-dependent gene products that we characterized, two small heat shock proteins, Hsp12 and Hsp122, were found to be involved in the polyene antifungal drug susceptibility of C. neoformans.
Figures

Similar articles
-
Noncanonical Gβ Gib2 is a scaffolding protein promoting cAMP signaling through functions of Ras1 and Cac1 proteins in Cryptococcus neoformans.J Biol Chem. 2014 May 2;289(18):12202-16. doi: 10.1074/jbc.M113.537183. Epub 2014 Mar 21. J Biol Chem. 2014. PMID: 24659785 Free PMC article.
-
Cross talk between the cell wall integrity and cyclic AMP/protein kinase A pathways in Cryptococcus neoformans.mBio. 2014 Aug 12;5(4):e01573-14. doi: 10.1128/mBio.01573-14. mBio. 2014. PMID: 25118241 Free PMC article.
-
Molecular Characterization of Adenylyl Cyclase Complex Proteins Using Versatile Protein-Tagging Plasmid Systems in Cryptococcus neoformans.J Microbiol Biotechnol. 2017 Feb 28;27(2):357-364. doi: 10.4014/jmb.1609.09036. J Microbiol Biotechnol. 2017. PMID: 27780958
-
The cAMP/Protein Kinase a Pathway Regulates Virulence and Adaptation to Host Conditions in Cryptococcus neoformans.Front Cell Infect Microbiol. 2019 Jun 18;9:212. doi: 10.3389/fcimb.2019.00212. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31275865 Free PMC article. Review.
-
Novel sensing mechanisms and targets for the cAMP-protein kinase A pathway in the yeast Saccharomyces cerevisiae.Mol Microbiol. 1999 Sep;33(5):904-18. doi: 10.1046/j.1365-2958.1999.01538.x. Mol Microbiol. 1999. PMID: 10476026 Review.
Cited by
-
Ras-Mediated Signal Transduction and Virulence in Human Pathogenic Fungi.Fungal Genom Biol. 2012;2(1):105. doi: 10.4172/2165-8056.1000105. Fungal Genom Biol. 2012. PMID: 24855584 Free PMC article.
-
Functional characterization of the small heat shock protein Hsp12p from Candida albicans.PLoS One. 2012;7(8):e42894. doi: 10.1371/journal.pone.0042894. Epub 2012 Aug 7. PLoS One. 2012. PMID: 22880130 Free PMC article.
-
The emerging fungal pathogen Cryptococcus gattii: Epidemiology, pathogenesis, immunomodulatory attributes, and drug susceptibility.PLoS Negl Trop Dis. 2025 Jul 3;19(7):e0013245. doi: 10.1371/journal.pntd.0013245. eCollection 2025 Jul. PLoS Negl Trop Dis. 2025. PMID: 40608829 Free PMC article. Review.
-
Genetic contribution to high temperature tolerance in Cryptococcus neoformans.Genetics. 2021 Mar 3;217(1):1-15. doi: 10.1093/genetics/iyaa009. Genetics. 2021. PMID: 33683363 Free PMC article.
-
Chaperone Networks in Fungal Pathogens of Humans.J Fungi (Basel). 2021 Mar 12;7(3):209. doi: 10.3390/jof7030209. J Fungi (Basel). 2021. PMID: 33809191 Free PMC article. Review.
References
-
- Alspaugh J. A., Cavallo L. M., Perfect J. R., Heitman J. 2000. RAS1 regulates filamentation, mating and growth at high temperature of Cryptococcus neoformans. Mol. Microbiol. 36:352–365 - PubMed
-
- Alspaugh J. A., Perfect J. R., Heitman J. 1998. Signal transduction pathways regulating differentiation and pathogenicity of Cryptococcus neoformans. Fungal Genet. Biol. 25:1–14 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
