

The neurosecretory vesicle protein phogrin functions as a phosphatidylinositol phosphatase to regulate insulin secretion
- PMID: 20097759
- PMCID: PMC2856256
- DOI: 10.1074/jbc.M109.066563
The neurosecretory vesicle protein phogrin functions as a phosphatidylinositol phosphatase to regulate insulin secretion
Abstract
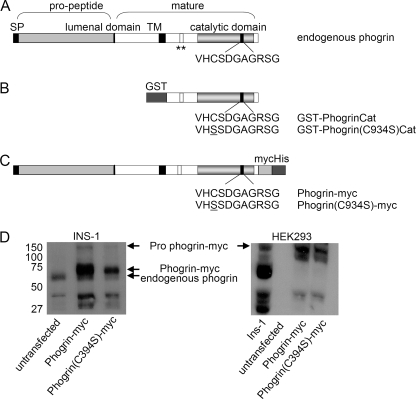
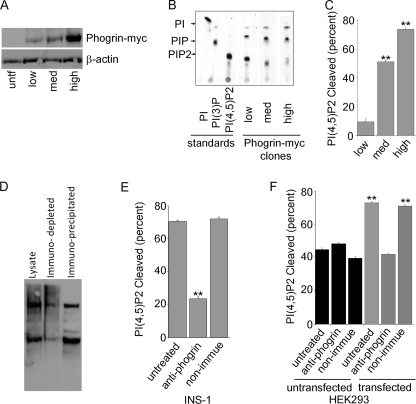
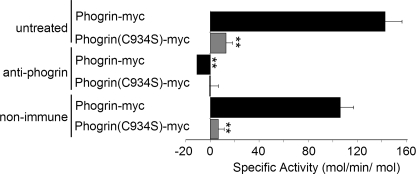
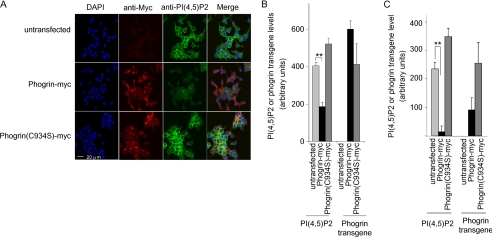
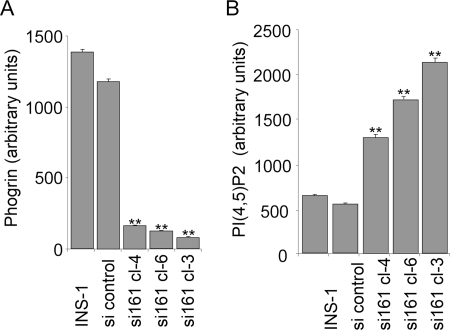
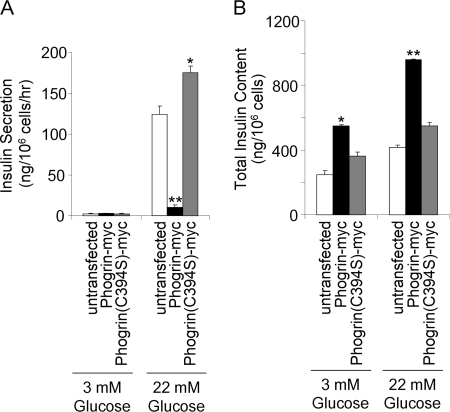
Phogrin is a transmembrane protein expressed in cells with stimulus-coupled peptide hormone secretion, including pancreatic beta cells, in which it is localized to the membrane of insulin-containing dense-core vesicles. By sequence, phogrin is a member of the family of receptor-like protein-tyrosine phosphatases, but it contains substitutions in conserved catalytic sequences, and no significant enzymatic activity for phogrin has ever been reported. We report here that phogrin is able to dephosphorylate specific inositol phospholipids, including phosphatidylinositol (PI) 3-phosphate and PI 4,5-diphosphate but not PI 3,4,5-trisphosphate. The phosphatidylinositol phosphatase (PIPase) activity of phogrin was measurable but low when evaluated by the ability of a catalytic domain fusion protein to hydrolyze soluble short-chain phosphatidylinositol phospholipids. Unlike most PIPases, which are cytoplasmic proteins that associate with membranes, mature phogrin is a transmembrane protein. When the transmembrane form of phogrin was overexpressed in mammalian cells, it reduced plasma membrane phosphatidylinositol 4,5-disphosphate levels in a dose-dependent manner. When purified and assayed in vitro, the transmembrane form had a specific activity of 142 mol/min/mol, 75-fold more active than the catalytic domain fusion protein and comparable with the specific activities of the other PIPases. The PIPase activity of phogrin depended on the catalytic site cysteine and correlated with effects on glucose-stimulated insulin secretion. We propose that phogrin functions as a phosphatidylinositol phosphatase that contributes to maintaining subcellular differences in levels of PIP that are important for regulating stimulus-coupled exocytosis of insulin.
Figures
Similar articles
-
Secretagogue-dependent phosphorylation of the insulin granule membrane protein phogrin is mediated by cAMP-dependent protein kinase.J Biol Chem. 2001 Aug 24;276(34):31919-28. doi: 10.1074/jbc.M102580200. Epub 2001 May 15. J Biol Chem. 2001. PMID: 11353772
-
Gene silencing of phogrin unveils its essential role in glucose-responsive pancreatic beta-cell growth.Diabetes. 2009 Mar;58(3):682-92. doi: 10.2337/db08-0970. Epub 2008 Dec 10. Diabetes. 2009. PMID: 19073770 Free PMC article.
-
The pseudophosphatase phogrin enables glucose-stimulated insulin signaling in pancreatic β cells.J Biol Chem. 2018 Apr 20;293(16):5920-5933. doi: 10.1074/jbc.RA117.000301. Epub 2018 Feb 26. J Biol Chem. 2018. PMID: 29483197 Free PMC article.
-
Insulin secretion by 'kiss-and-run' exocytosis in clonal pancreatic islet beta-cells.Biochem Soc Trans. 2003 Aug;31(Pt 4):833-6. doi: 10.1042/bst0310833. Biochem Soc Trans. 2003. PMID: 12887316 Review.
-
Phosphatidylinositol Kinases and Phosphatases in Entamoeba histolytica.Front Cell Infect Microbiol. 2019 Jun 6;9:150. doi: 10.3389/fcimb.2019.00150. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31245297 Free PMC article. Review.
Cited by
-
Label-free Quantitative Proteomics of Mouse Cerebrospinal Fluid Detects β-Site APP Cleaving Enzyme (BACE1) Protease Substrates In Vivo.Mol Cell Proteomics. 2015 Oct;14(10):2550-63. doi: 10.1074/mcp.M114.041533. Epub 2015 Jul 2. Mol Cell Proteomics. 2015. PMID: 26139848 Free PMC article.
-
The Ia-2β intronic miRNA, miR-153, is a negative regulator of insulin and dopamine secretion through its effect on the Cacna1c gene in mice.Diabetologia. 2015 Oct;58(10):2298-306. doi: 10.1007/s00125-015-3683-8. Epub 2015 Jul 4. Diabetologia. 2015. PMID: 26141787 Free PMC article.
-
Co-regulation of intragenic microRNA miR-153 and its host gene Ia-2 β: identification of miR-153 target genes with functions related to IA-2β in pancreas and brain.Diabetologia. 2013 Jul;56(7):1547-56. doi: 10.1007/s00125-013-2901-5. Epub 2013 Apr 18. Diabetologia. 2013. PMID: 23595248 Free PMC article.
-
Role of adaptor proteins in secretory granule biogenesis and maturation.Front Endocrinol (Lausanne). 2013 Aug 14;4:101. doi: 10.3389/fendo.2013.00101. eCollection 2013. Front Endocrinol (Lausanne). 2013. PMID: 23966980 Free PMC article.
-
Genome-wide associations between genetic and epigenetic variation influence mRNA expression and insulin secretion in human pancreatic islets.PLoS Genet. 2014 Nov 6;10(11):e1004735. doi: 10.1371/journal.pgen.1004735. eCollection 2014 Nov. PLoS Genet. 2014. PMID: 25375650 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
