

Improved strains and plasmid vectors for conditional overexpression of His-tagged proteins in Haloferax volcanii
- PMID: 20097827
- PMCID: PMC2838008
- DOI: 10.1128/AEM.02670-09
Improved strains and plasmid vectors for conditional overexpression of His-tagged proteins in Haloferax volcanii
Abstract
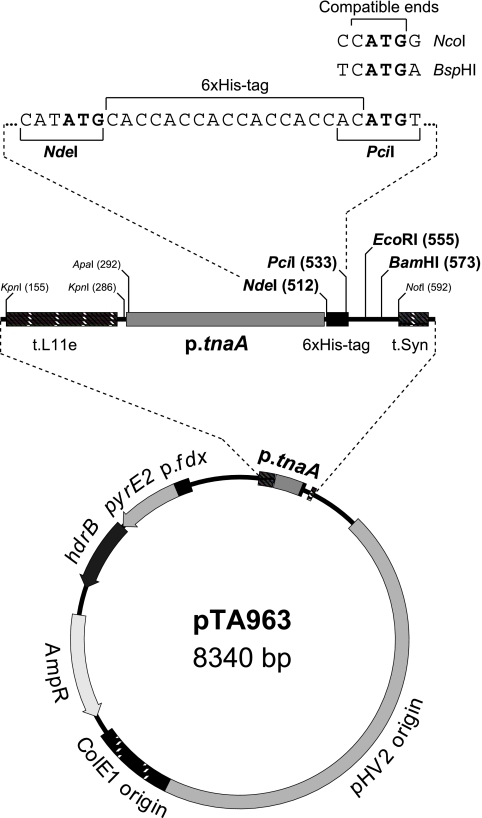
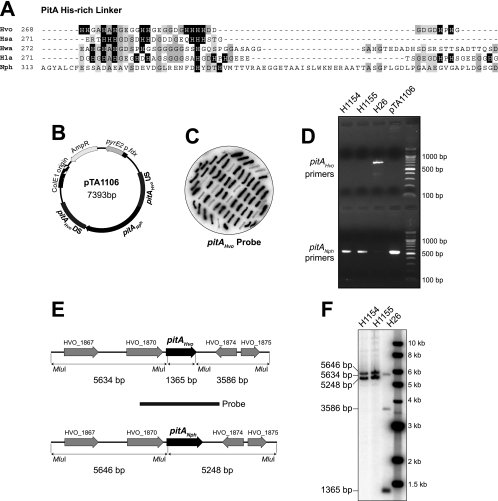
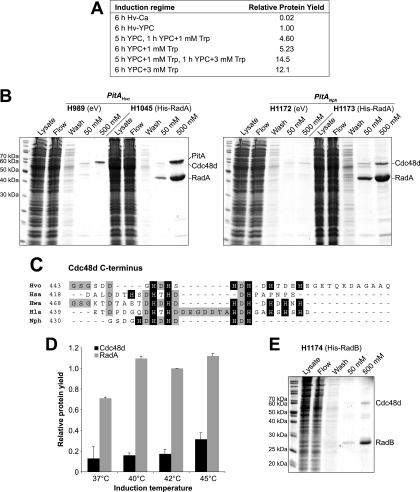
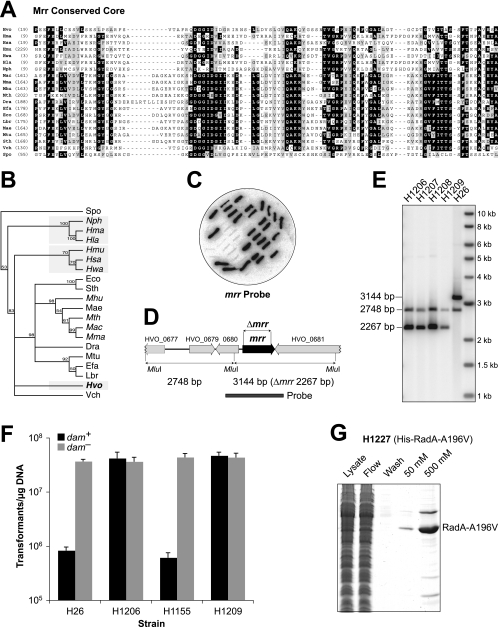
Research into archaea will not achieve its full potential until systems are in place to carry out genetics and biochemistry in the same species. Haloferax volcanii is widely regarded as the best-equipped organism for archaeal genetics, but the development of tools for the expression and purification of H. volcanii proteins has been neglected. We have developed a series of plasmid vectors and host strains for conditional overexpression of halophilic proteins in H. volcanii. The plasmids feature the tryptophan-inducible p.tnaA promoter and a 6xHis tag for protein purification by metal affinity chromatography. Purification is facilitated by host strains, where pitA is replaced by the ortholog from Natronomonas pharaonis. The latter lacks the histidine-rich linker region found in H. volcanii PitA and does not copurify with His-tagged recombinant proteins. We also deleted the mrr restriction endonuclease gene, thereby allowing direct transformation without the need to passage DNA through an Escherichia coli dam mutant.
Figures
References
-
- Aertsen, A., M. Tesfazgi Mebrhatu, and C. W. Michiels. 2008. Activation of the Salmonella typhimurium Mrr protein. Biochem. Biophys. Res. Commun. 367:435-439. - PubMed
-
- Allers, T., and M. Mevarech. 2005. Archaeal genetics—the third way. Nat. Rev. Genet. 6:58-73. - PubMed
-
- Bab-Dinitz, E., H. Shmuely, J. Maupin-Furlow, J. Eichler, and B. Shaanan. 2006. Haloferax volcanii PitA: an example of functional interaction between the Pfam chlorite dismutase and antibiotic biosynthesis monooxygenase families? Bioinformatics 22:671-675. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
