

Acute regulation of the epithelial sodium channel in airway epithelia by proteases and trafficking
- PMID: 20097829
- PMCID: PMC2993091
- DOI: 10.1165/rcmb.2009-0348OC
Acute regulation of the epithelial sodium channel in airway epithelia by proteases and trafficking
Abstract
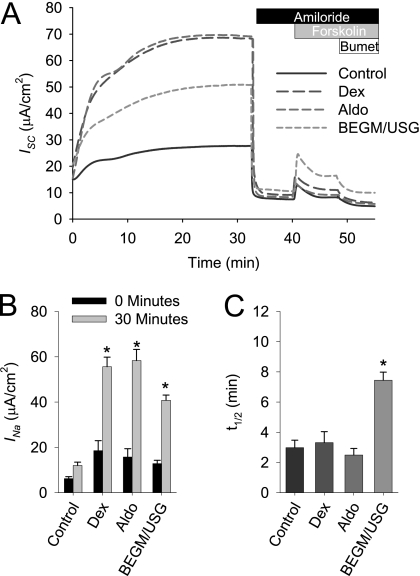
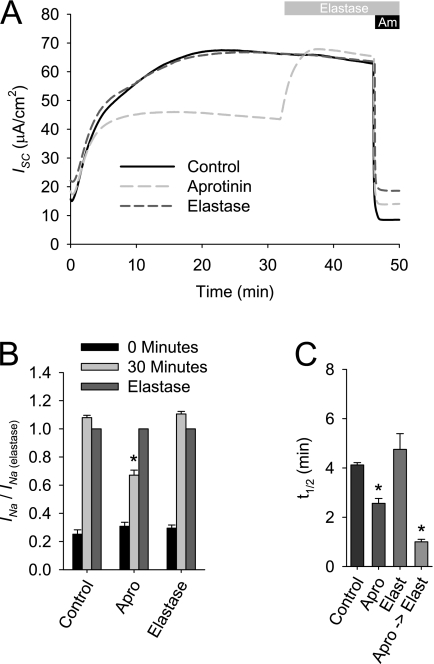
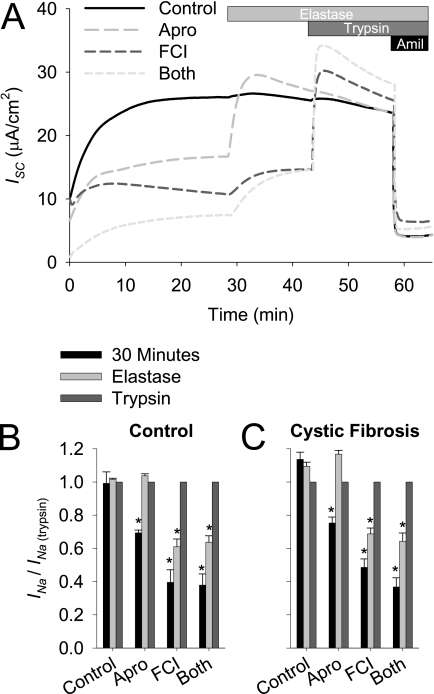
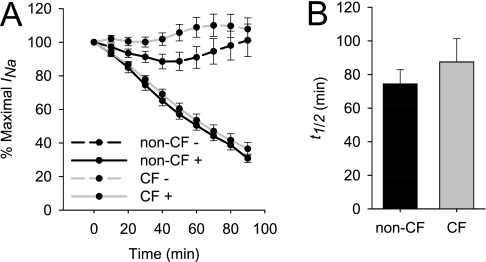
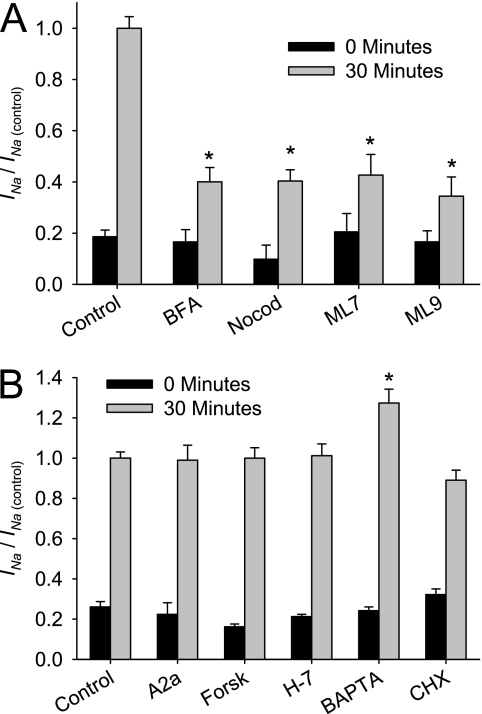
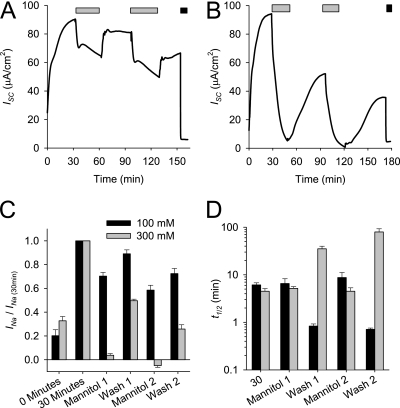
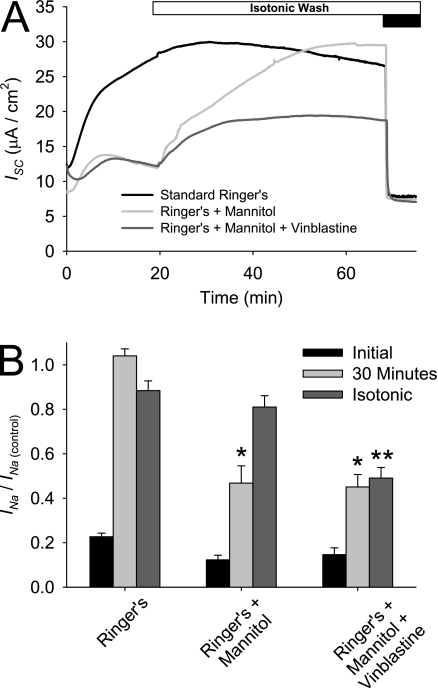
Effective clearance of inhaled pathogens is the primary innate defense mechanism in the lung, and requires the maintenance of a proper airway surface liquid (ASL) volume to facilitate ciliary beat and optimize mucociliary clearance. Na(+) absorption via the epithelial sodium channel (ENaC) is tightly regulated and, together with chloride movement, provides the optimal osmotic gradients to absorb excessive fluid in the airway lumen while preventing excessive ASL dehydration, which would compromise mucus clearance from the lung. To absorb excessive fluid from the luminal surface, a local mechanism of ENaC activation allows for an increase in Na(+) absorption at times when the ASL volume is expanded. To help define these regulatory mechanisms, we examined the effects of ASL volume expansion on ENaC activity in primary human bronchial epithelial (HBE) cell cultures. We found that ENaC activity increases dramatically after rapid dilution of endogenous ASL. Approximately 35% of the increase in Na(+) absorption was attributable to activation of ENaC by proteases. The remainder of the increase in Na(+) current was prevented when membrane trafficking was disrupted with brefeldin A, nocodazole, or myosin light chain kinase inhibitors, demonstrating that trafficking is involved with ENaC regulation in the airway. These findings demonstrate that Na(+) absorption in the airway is acutely modulated by the coordinated trafficking of channels to the luminal surface and by the proteolytic activation of ENaC in response to ASL volume expansion.
Figures
References
-
- Matsui H, Grubb BR, Tarran R, Randell SH, Gatzy JT, Davis CW, Boucher RC. Evidence for periciliary liquid layer depletion, not abnormal ion composition, in the pathogenesis of cystic fibrosis airways disease. Cell 1998;95:1005–1015. - PubMed
-
- Donaldson SH, Boucher RC. Sodium channels and cystic fibrosis. Chest 2007;132:1631–1636. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
