

Direct evidence for control of the pheromone-inducible prgQ operon of Enterococcus faecalis plasmid pCF10 by a countertranscript-driven attenuation mechanism
- PMID: 20097859
- PMCID: PMC2832536
- DOI: 10.1128/JB.01525-09
Direct evidence for control of the pheromone-inducible prgQ operon of Enterococcus faecalis plasmid pCF10 by a countertranscript-driven attenuation mechanism
Abstract
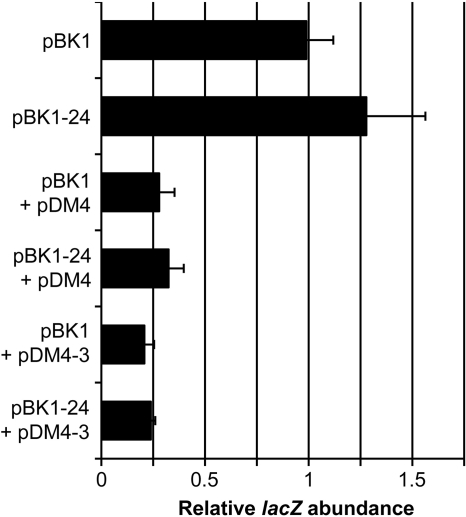
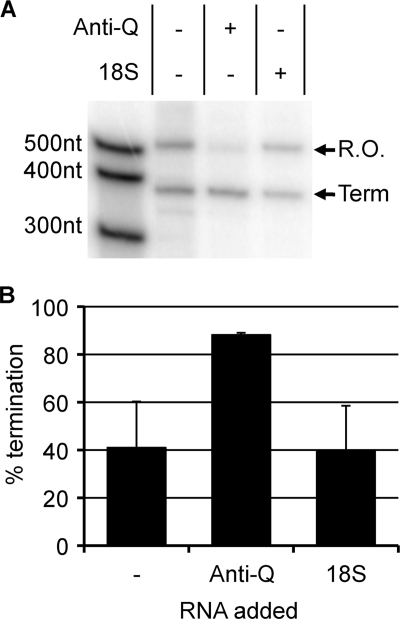
The mating response of Enterococcus faecalis cells carrying the conjugative plasmid pCF10 is controlled by multiple regulatory circuits. Initiation of transcription of the prgQ conjugation operon is controlled by the peptide receptor protein PrgX; binding of the pheromone peptide cCF10 to PrgX abolishes PrgX repression, while binding of the inhibitor peptide iCF10 enhances repression. The results of molecular analysis of prgQ transcripts and genetic studies suggested that the elongation of prgQ transcripts past a putative terminator (IRS1) may be controlled by the interaction of nascent prgQ mRNAs with a small antisense RNA (Anti-Q) encoded within prgQ. Direct evidence for interaction of these RNAs, as well as the resulting effects on readthrough of prgQ transcription, has been limited. Here we report the results of experiments that (i) determine the inherent termination properties of prgQ transcripts in the absence of Anti-Q; (ii) determine the direct effects of the interaction of Anti-Q with nascent prgQ transcripts in the absence of complicating effects of the PrgX protein; and (iii) begin to dissect the structural components involved in these interactions. The results confirm the existence of alternative terminating and antiterminating forms of nascent prgQ transcripts in vivo and demonstrate that the interaction of Anti-Q with these transcripts leads to termination via inhibition of antiterminator formation. In vitro transcription assays support the major results of the in vivo studies. The data support a model for Anti-Q function suggested from recent studies of these RNAs and their interactions in vitro (S. Shokeen, C. M. Johnson, T. J. Greenfield, D. A. Manias, G. M. Dunny, and K. E. Weaver, submitted for publication).
Figures


Similar articles
-
Enterococcal Sex Pheromones: Evolutionary Pathways to Complex, Two-Signal Systems.J Bacteriol. 2016 May 13;198(11):1556-1562. doi: 10.1128/JB.00128-16. Print 2016 Jun 1. J Bacteriol. 2016. PMID: 27021562 Free PMC article. Review.
-
In vivo and in vitro analyses of regulation of the pheromone-responsive prgQ promoter by the PrgX pheromone receptor protein.J Bacteriol. 2012 Jul;194(13):3386-94. doi: 10.1128/JB.00364-12. Epub 2012 Apr 27. J Bacteriol. 2012. PMID: 22544272 Free PMC article.
-
Characterization of cis-acting prgQ mutants: evidence for two distinct repression mechanisms by Qa RNA and PrgX protein in pheromone-inducible enterococcal plasmid pCF10.Mol Microbiol. 2004 Jan;51(1):271-81. doi: 10.1046/j.1365-2958.2003.03832.x. Mol Microbiol. 2004. PMID: 14651627
-
Analysis of expression of prgX, a key negative regulator of the transfer of the Enterococcus faecalis pheromone-inducible plasmid pCF10.J Mol Biol. 2000 Apr 7;297(4):861-75. doi: 10.1006/jmbi.2000.3628. J Mol Biol. 2000. PMID: 10736223
-
The peptide pheromone-inducible conjugation system of Enterococcus faecalis plasmid pCF10: cell-cell signalling, gene transfer, complexity and evolution.Philos Trans R Soc Lond B Biol Sci. 2007 Jul 29;362(1483):1185-93. doi: 10.1098/rstb.2007.2043. Philos Trans R Soc Lond B Biol Sci. 2007. PMID: 17360276 Free PMC article. Review.
Cited by
-
MODELING OF GENE REGULATORY PROCESSES BY POPULATION MEDIATED SIGNALING. NEW APPLICATIONS OF POPULATION BALANCES.Chem Eng Sci. 2012 Mar 5;70:188-199. doi: 10.1016/j.ces.2011.07.062. Epub 2011 Aug 11. Chem Eng Sci. 2012. PMID: 22581980 Free PMC article.
-
Structure and mode of peptide binding of pheromone receptor PrgZ.J Biol Chem. 2012 Oct 26;287(44):37165-70. doi: 10.1074/jbc.M112.386334. Epub 2012 Sep 4. J Biol Chem. 2012. PMID: 22948145 Free PMC article.
-
Examination of Enterococcus faecalis Toxin-Antitoxin System Toxin Fst Function Utilizing a Pheromone-Inducible Expression Vector with Tight Repression and Broad Dynamic Range.J Bacteriol. 2017 May 25;199(12):e00065-17. doi: 10.1128/JB.00065-17. Print 2017 Jun 15. J Bacteriol. 2017. PMID: 28348028 Free PMC article.
-
Enterococcal Sex Pheromones: Evolutionary Pathways to Complex, Two-Signal Systems.J Bacteriol. 2016 May 13;198(11):1556-1562. doi: 10.1128/JB.00128-16. Print 2016 Jun 1. J Bacteriol. 2016. PMID: 27021562 Free PMC article. Review.
-
Characterization of the effects of an rpoC mutation that confers resistance to the Fst peptide toxin-antitoxin system toxin.J Bacteriol. 2013 Jan;195(1):156-66. doi: 10.1128/JB.01597-12. Epub 2012 Oct 26. J Bacteriol. 2013. PMID: 23104812 Free PMC article.
References
-
- Bae, T., S. Clerc-Bardin, and G. M. Dunny. 2000. Analysis of expression of prgX, a key negative regulator of the transfer of the Enterococcus faecalis pheromone-inducible plasmid pCF10. J. Mol. Biol. 297:861-875. - PubMed
-
- Bae, T., B. K. Kozlowicz, and G. M. Dunny. 2002. Two targets in pCF10 DNA for PrgX binding: their role in production of Anti-Q and prgX mRNA and in regulation of pheromone-inducible conjugation. J. Mol. Biol. 315:995-1007. - PubMed
-
- Bae, T., B. K. Kozlowicz, and G. M. Dunny. 2004. Characterization of cis-acting prgQ mutants: evidence for two distinct repression mechanisms by Anti-Q RNA and PrgX protein in pheromone-inducible enterococcal plasmid pCF10. Mol. Microbiol. 51:271-281. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
