

Differential effector pathways regulate memory CD8 T cell immunity against Plasmodium berghei versus P. yoelii sporozoites
- PMID: 20097864
- PMCID: PMC2904689
- DOI: 10.4049/jimmunol.0903529
Differential effector pathways regulate memory CD8 T cell immunity against Plasmodium berghei versus P. yoelii sporozoites
Abstract
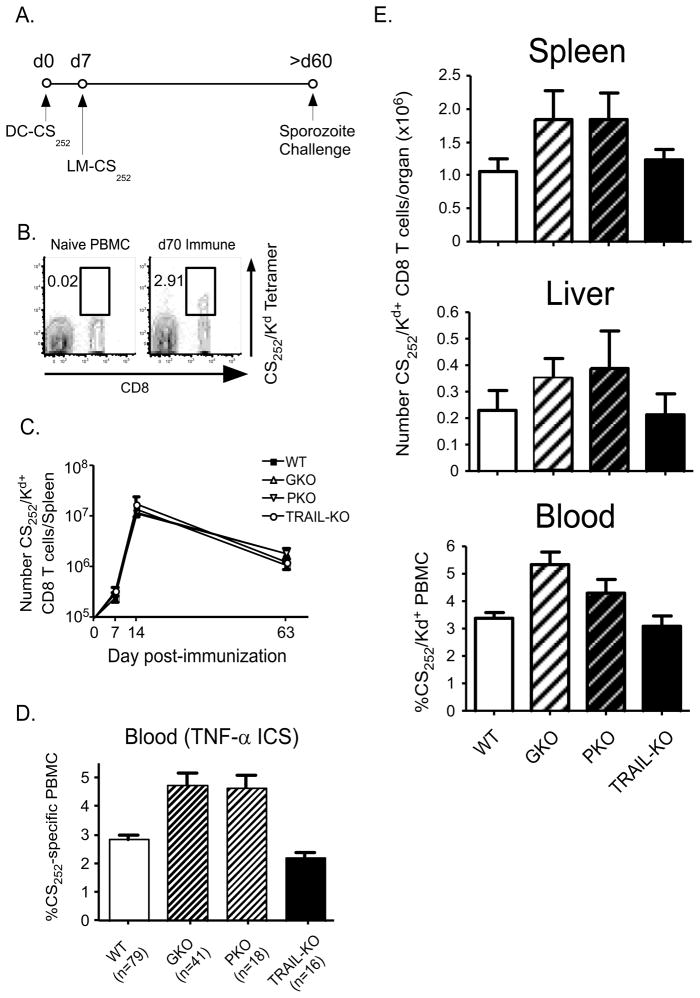
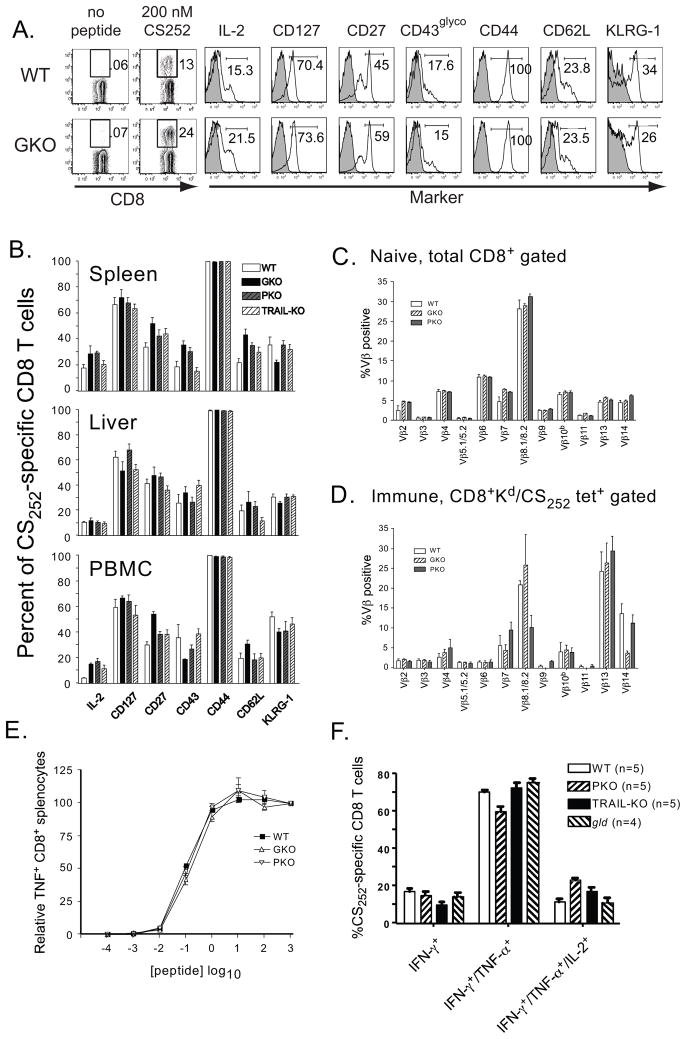
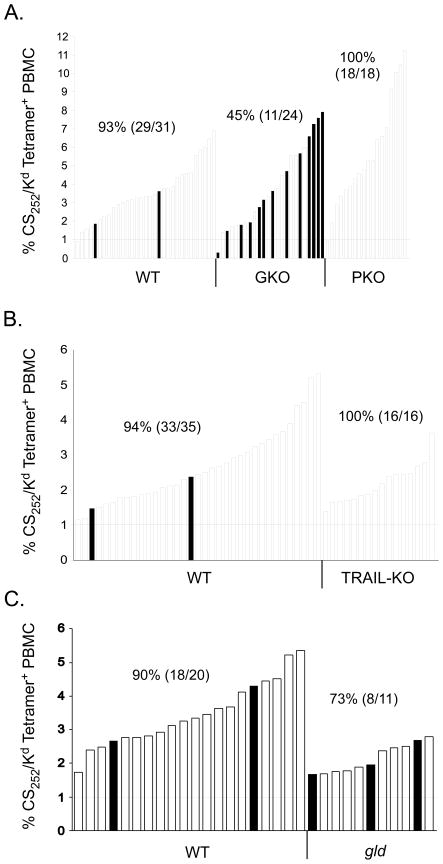
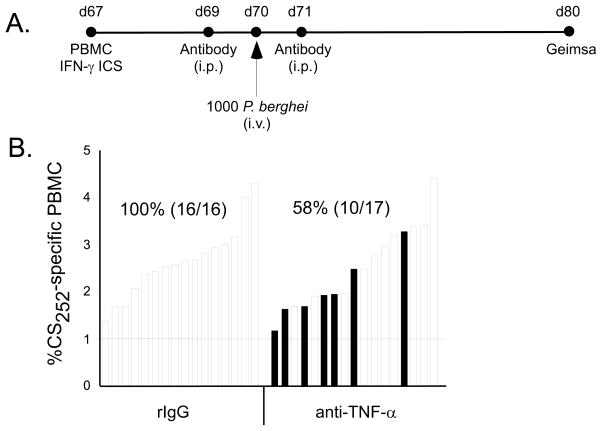
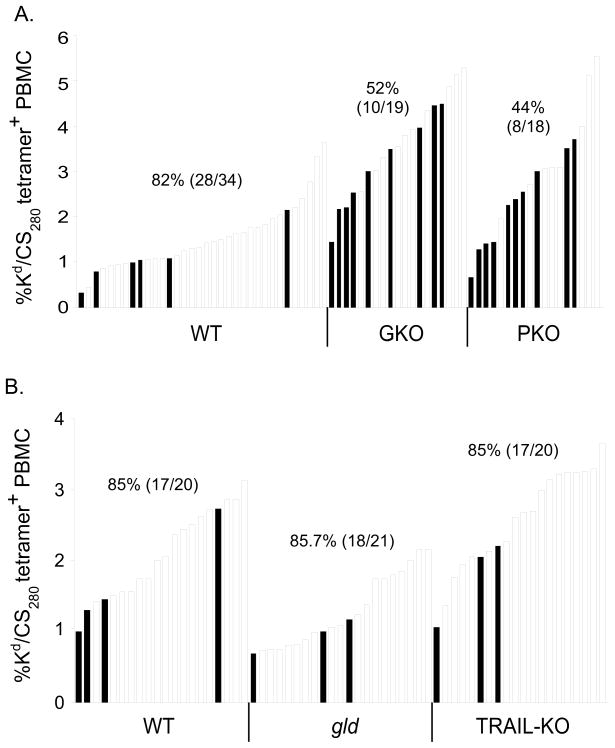
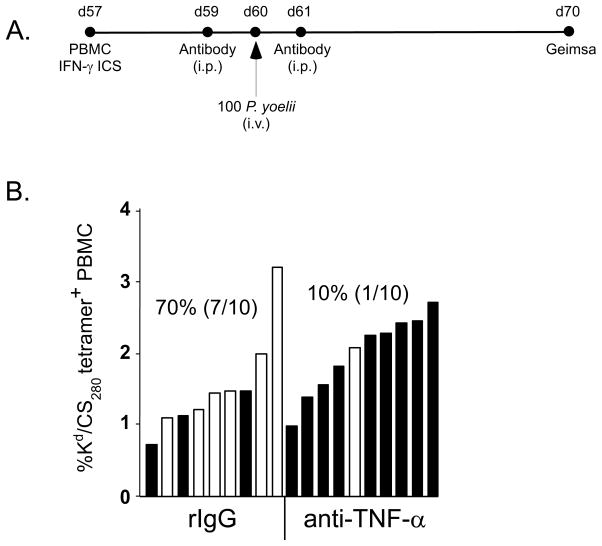
Malaria results in >1,000,000 deaths per year worldwide. Although no licensed vaccine exists, much effort is currently focused on subunit vaccines that elicit CD8 T cell responses directed against Plasmodium parasite liver stage Ags. Multiple immune-effector molecules play a role in antimicrobial immunity mediated by memory CD8 T cells, including IFN-gamma, perforin, TRAIL, Fas ligand, and TNF-alpha. However, it is not known which pathways are required for memory CD8 T cell-mediated immunity against liver stage Plasmodium infection. In this study, we used a novel immunization strategy to generate memory CD8 T cells in the BALB/c mouse model of P. berghei or P. yoelii sporozoite infection to examine the role of immune-effector molecules in resistance to the liver stage infection. Our studies reveal that endogenous memory CD8 T cell-mediated protection against both parasite species is, in part, dependent on IFN-gamma, whereas perforin was only critical in protection against P. yoelii. We further show that neutralization of TNF-alpha in immunized mice markedly reduces memory CD8 T cell-mediated protection against both parasite species. Thus, our studies identify IFN-gamma and TNF-alpha as important components of the noncytolytic pathways that underlie memory CD8 T cell-mediated immunity against liver stage Plasmodium infection. Our studies also show that the effector pathways that memory CD8 T cells use to eliminate liver stage infection are, in part, Plasmodium species specific.
Figures

References
-
- Todryk SM, Hill AV. Malaria vaccines: the stage we are at. Nature reviews. 2007;5:487–489. - PubMed
-
- Bryce J, Boschi-Pinto C, Shibuya K, Black RE. WHO estimates of the causes of death in children. Lancet. 2005;365:1147–1152. - PubMed
-
- Sidjanski S, Vanderberg JP. Delayed migration of Plasmodium sporozoites from the mosquito bite site to the blood. The American journal of tropical medicine and hygiene. 1997;57:426–429. - PubMed
-
- Amino R, Thiberge S, Martin B, Celli S, Shorte S, Frischknecht F, Menard R. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nature medicine. 2006;12:220–224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
