

DNA zip codes control an ancient mechanism for gene targeting to the nuclear periphery
- PMID: 20098417
- PMCID: PMC2835469
- DOI: 10.1038/ncb2011
DNA zip codes control an ancient mechanism for gene targeting to the nuclear periphery
Erratum in
- Nat Cell Biol. 2010 Mar;12(3):306
Abstract
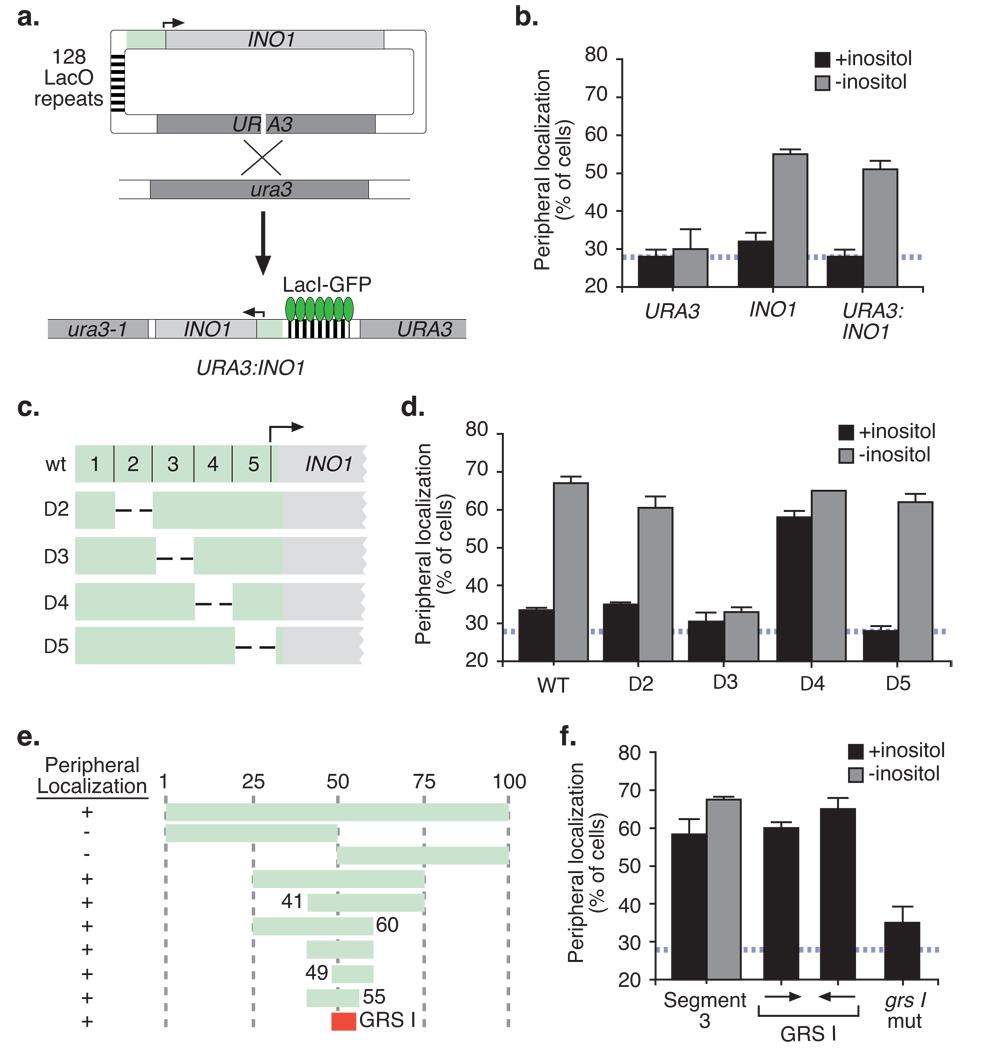
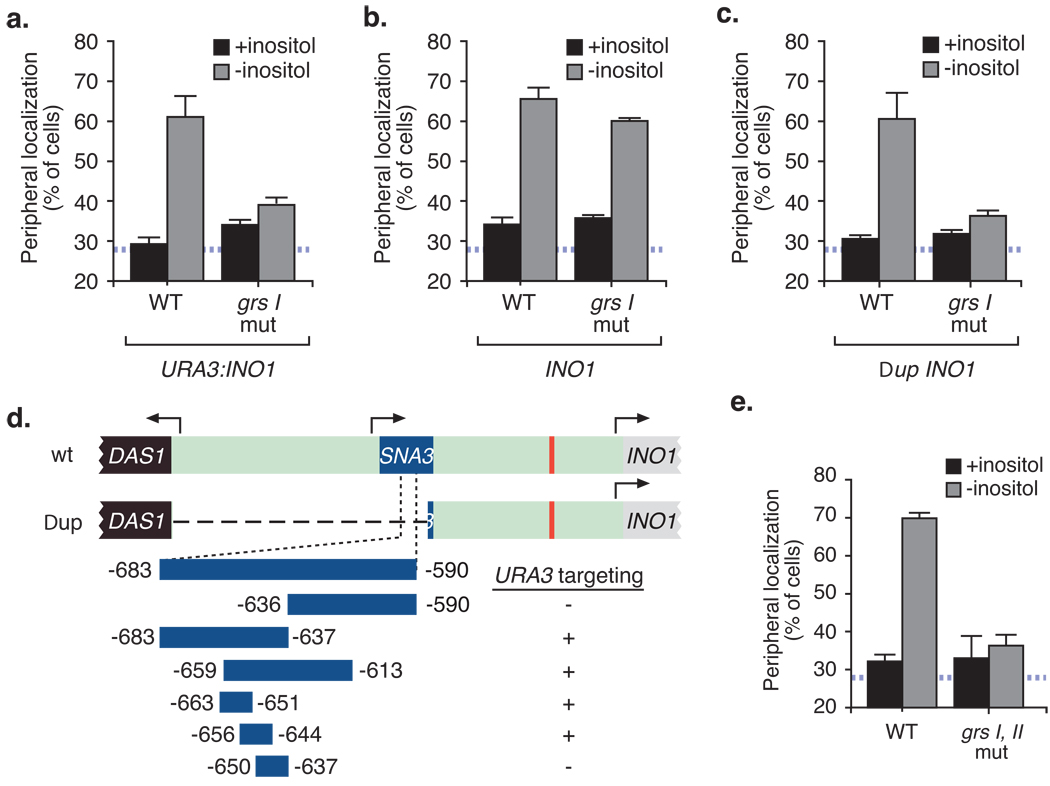
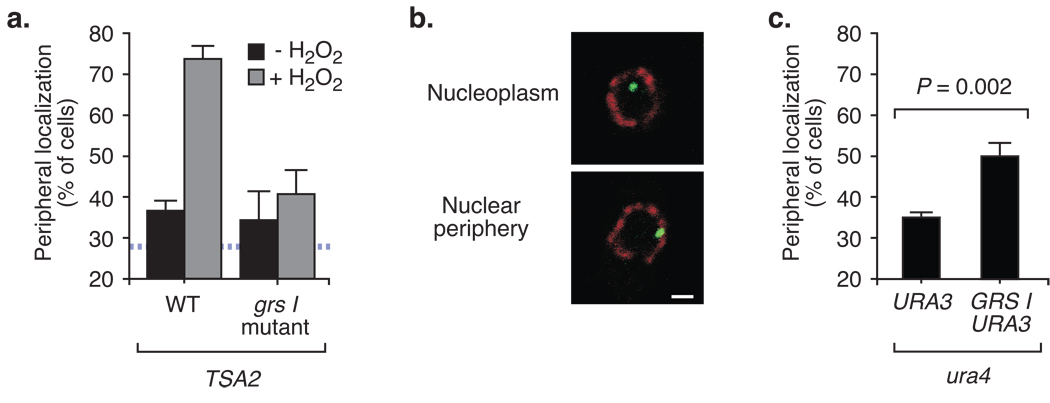
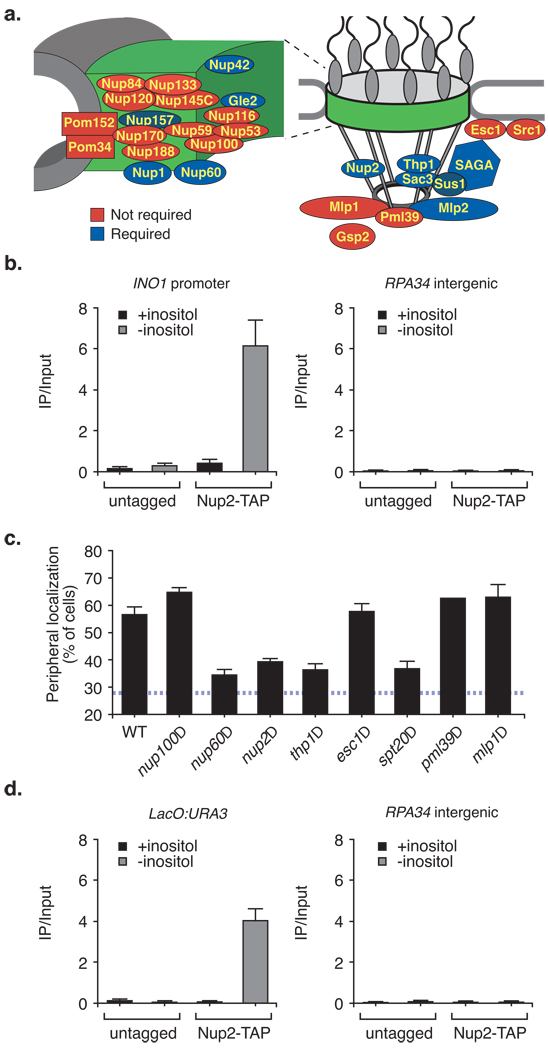
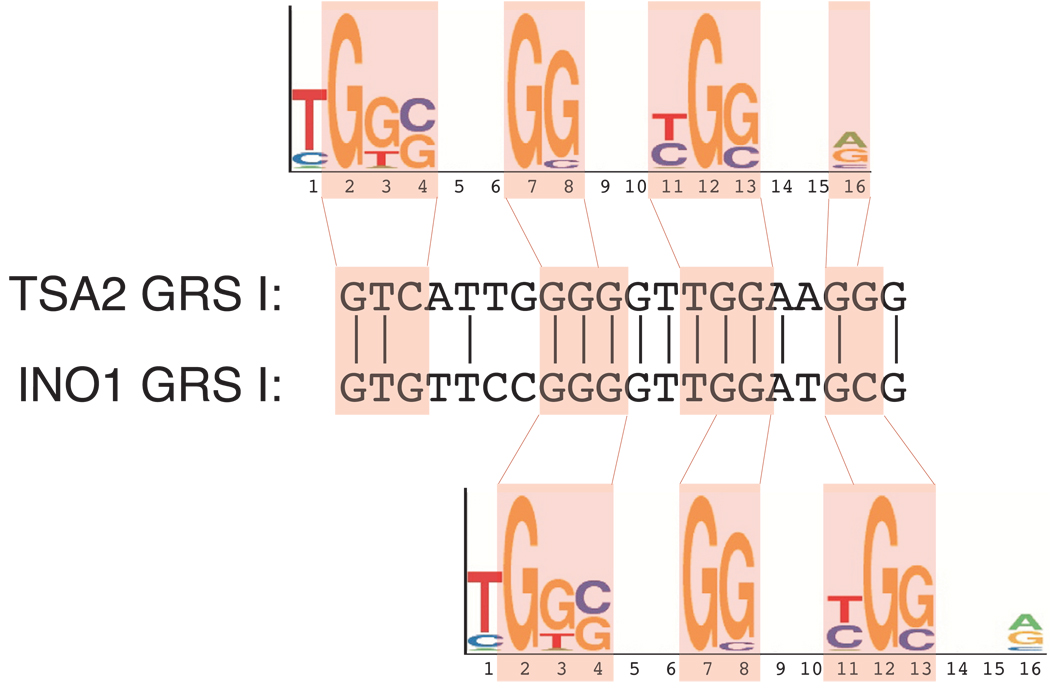
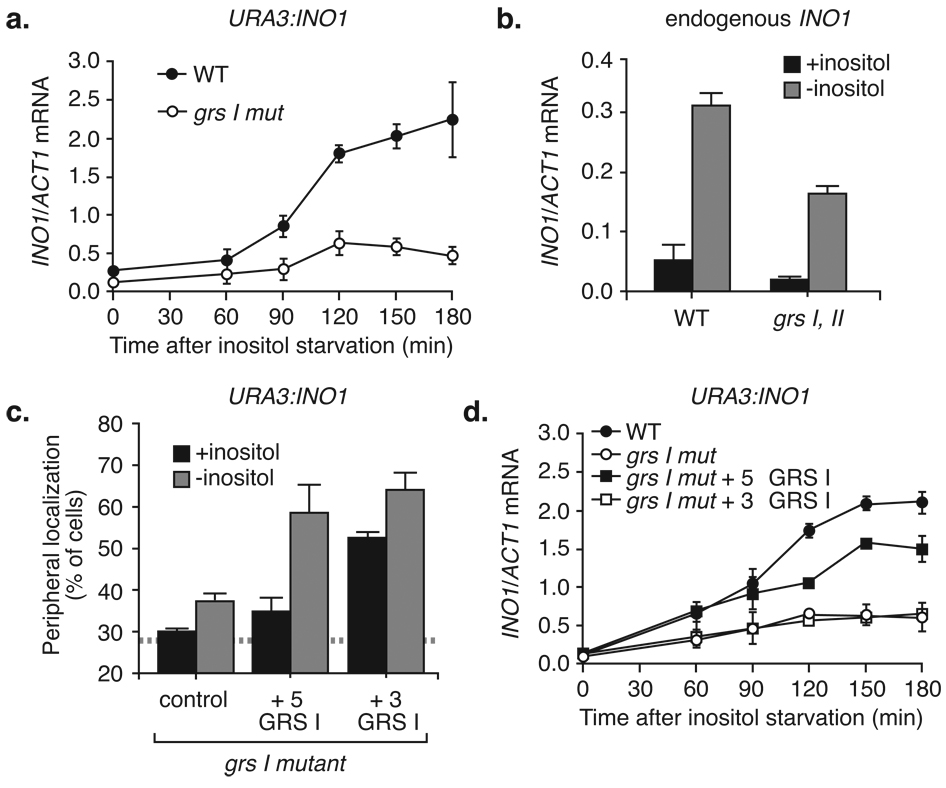
Many genes in Saccharomyces cerevisiae are recruited to the nuclear periphery after transcriptional activation. We have identified two gene recruitment sequences (GRS I and II) from the promoter of the INO1 gene that target the gene to the nuclear periphery. These GRSs function as DNA zip codes and are sufficient to target a nucleoplasmic locus to the nuclear periphery. Targeting requires components of the nuclear pore complex (NPC) and a GRS is sufficient to confer a physical interaction with the NPC. GRS I elements are enriched in promoters of genes that interact with the NPC, and genes that are induced by protein folding stress. Full transcriptional activation of INO1 and another GRS-containing gene requires GRS-mediated targeting of the promoter to the nuclear periphery. Finally, GRS I also functions as a DNA zip code in Schizosaccharomyces pombe, suggesting that this mechanism of targeting to the nuclear periphery has been conserved over approximately one billion years of evolution.
Figures
References
-
- Akhtar A, Gasser SM. The nuclear envelope and transcriptional control. Nat Rev Genet. 2007;8:507–517. - PubMed
-
- Ahmed S, Brickner JH. Regulation and epigenetic control of transcription at the nuclear periphery. Trends Genet. 2007;23:396–402. - PubMed
-
- Taddei A. Active genes at the nuclear pore complex. Curr Opin Cell Biol. 2007;19:305–310. - PubMed
-
- Casolari JM, et al. Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell. 2004;117:427–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
