

Rapid, nondenaturing RNA purification using weak anion-exchange fast performance liquid chromatography
- PMID: 20100812
- PMCID: PMC2822929
- DOI: 10.1261/rna.1862210
Rapid, nondenaturing RNA purification using weak anion-exchange fast performance liquid chromatography
Abstract
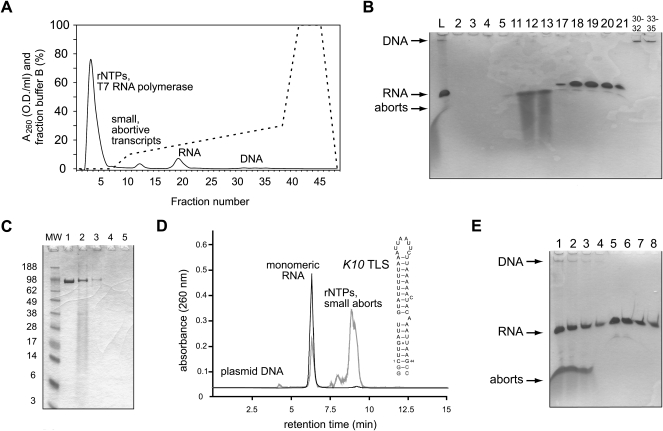
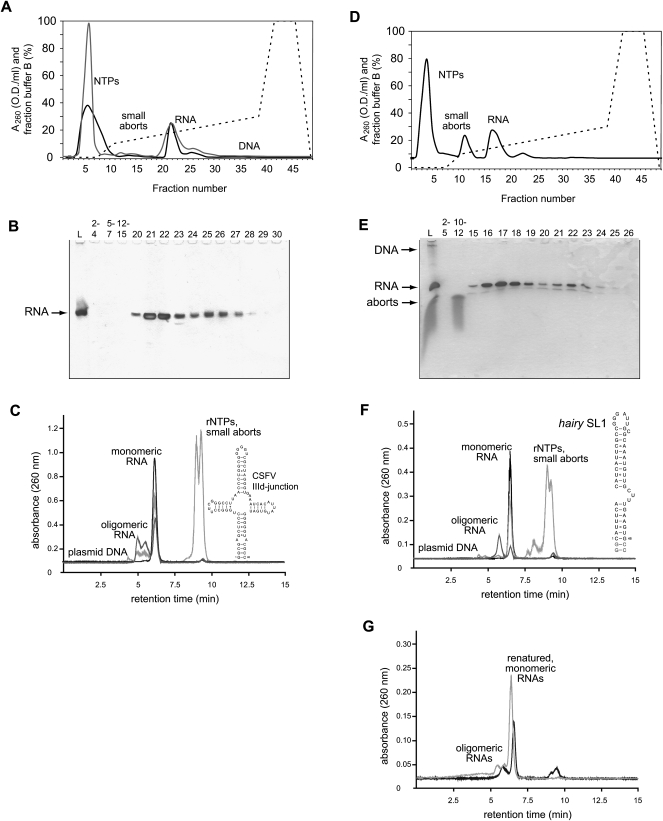
We present a simple and fast method for large-scale purification of RNA oligonucleotides suitable for biochemical and structural studies. RNAs are transcribed in vitro with T7 RNA polymerase using linearized plasmid DNA templates. After addition of EDTA, the crude transcription reaction is subjected directly to weak anion-exchange chromatography using DEAE-sepharose to separate the T7 RNA polymerase, unincorporated rNTPs, small abortive transcripts, and the plasmid DNA template from the desired RNA product. The novel method does neither require tedious phenol/chloroform extraction of the T7 RNA polymerase nor denaturation of the RNA, which is desirable especially for larger RNAs. In addition, isotopically labeled rNTPs can be easily recycled from the column flow-through and oligomeric RNA aggregates can be separated from the natively folded monomeric RNA product.
Figures

References
-
- Doudna JA. Preparation of homogeneous ribozyme RNA for crystallization. Methods Mol Biol. 1997;74:365–370. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources