

Intracellular MHC class II controls regulatory tolerance to allogeneic transplants
- PMID: 20100928
- PMCID: PMC2881546
- DOI: 10.4049/jimmunol.0803664
Intracellular MHC class II controls regulatory tolerance to allogeneic transplants
Abstract
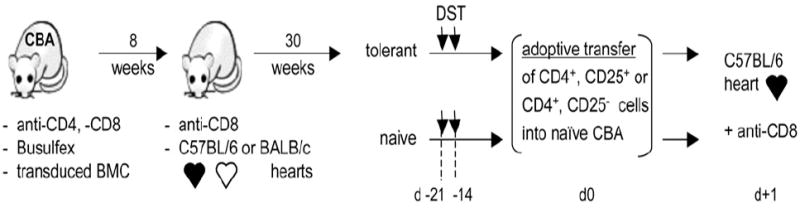
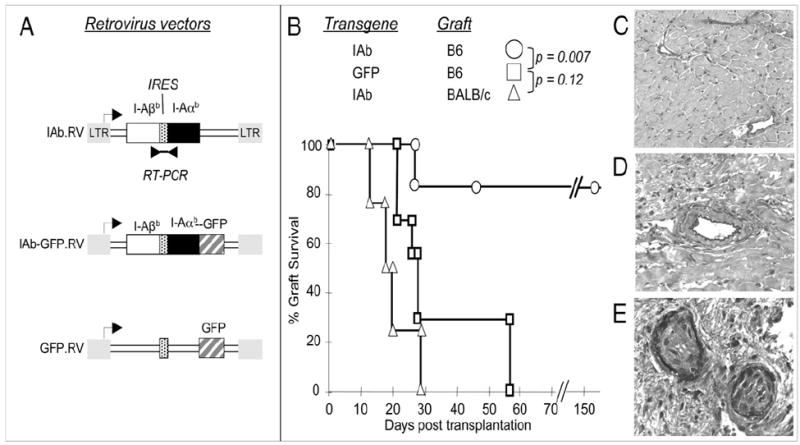
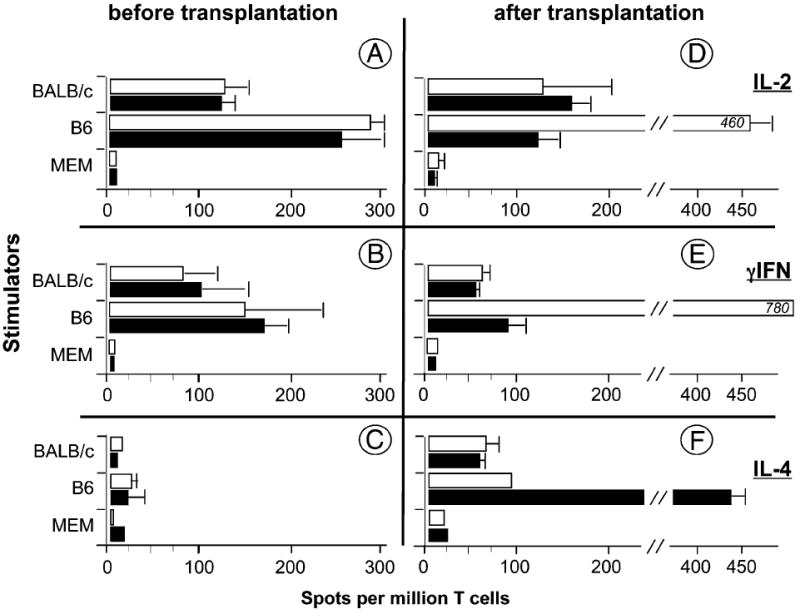
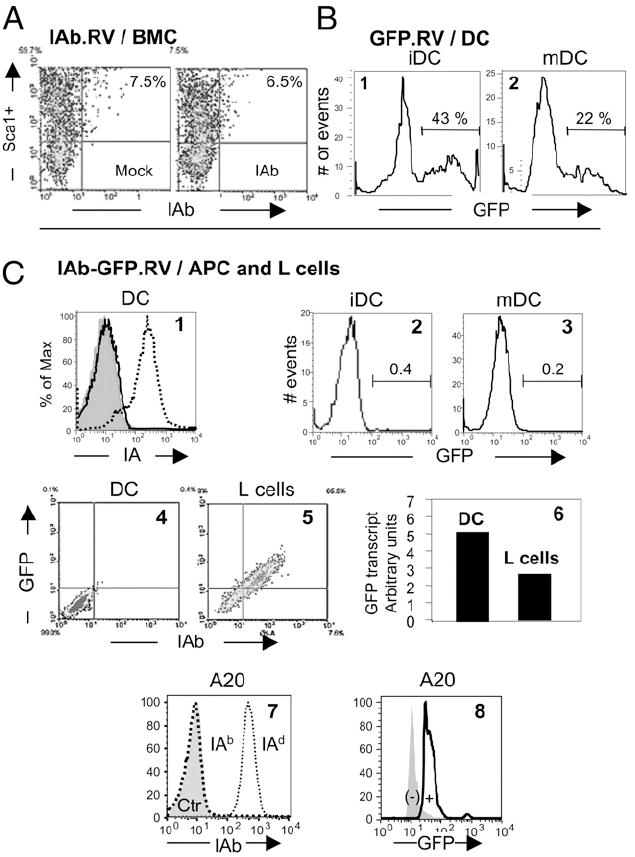
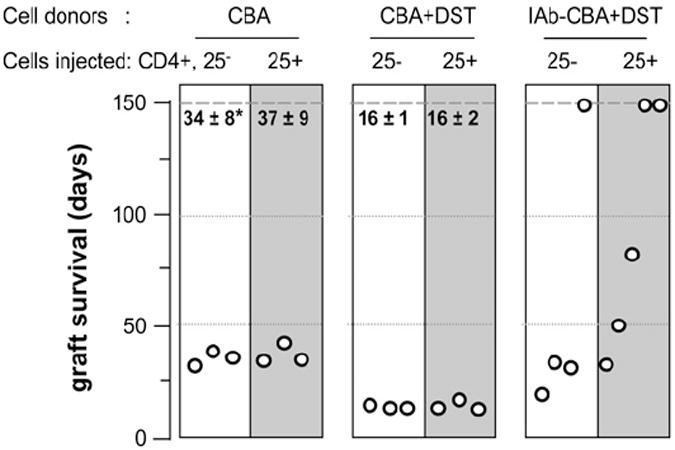
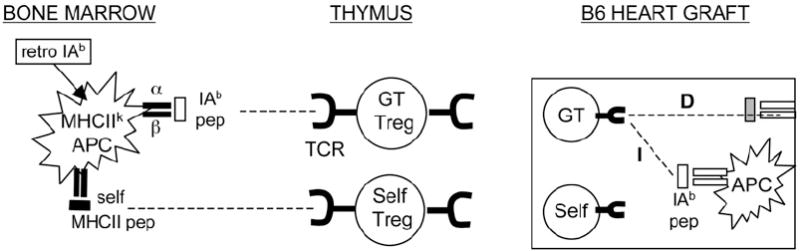
MHC class II (MHCII) genes have been implicated in the regulation of T lymphocyte responses. However, the mechanism of MHCII-driven regulation remains unknown. Matching for MHCII between donors and recipients of allografts favors regulatory T cell tolerance to transplants and provides a unique opportunity to study this regulation. In this study, we investigated MHCII regulation using transfer of donor MHCII genes in recipients of cardiac allografts. Transfer of MHCII IA(b) genes in the bone marrow of CBA mice (H-2(k)) prior to the grafting of IA(b+) fully allogeneic C57BL/6 (B6, H-2(b)) heart transplants resulted in donor-specific tolerance associated with long-term survival of B6, but not third-party, allografts without sustained immunosuppression. Strikingly, the majority of accepted heart transplants (>170 d) were devoid of allograft vasculopathy. Further studies indicated that intracellular IA(b) initiated the tolerogenic process, which was mediated by regulatory T cells (Tregs) that polarized antigraft responses to Th2 cytokine producers. This mechanism seems to be unique to MHCII genes, because previous MHC class I gene-based therapies failed to produce Tregs. These results demonstrate the key role of MHCII in the induction of Tregs. They also underscore a potential mechanism of specific inactivation of T cells in this model; when activated by IA(b+) grafts, IA(b)-specific Tregs repress the entire alloresponse to C57BL/6 transplants (including MHC I and minor Ags), thus mediating T cell tolerance.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures

Similar articles
-
Conferring indirect allospecificity on CD4+CD25+ Tregs by TCR gene transfer favors transplantation tolerance in mice.J Clin Invest. 2008 Nov;118(11):3619-28. doi: 10.1172/JCI33185. Epub 2008 Oct 9. J Clin Invest. 2008. PMID: 18846251 Free PMC article.
-
Intrathymic tolerance induction: determination of tolerance to class II major histocompatibility complex antigens in maturing T lymphocytes by a bone marrow-derived non-lymphoid thymus cell.Scand J Immunol. 1987 Dec;26(6):589-601. doi: 10.1111/j.1365-3083.1987.tb02294.x. Scand J Immunol. 1987. PMID: 3501157
-
Retroviral transfer of donor MHC class I or MHC class II genes into recipient bone marrow cells can induce operational tolerance to alloantigens in vivo.Hum Gene Ther. 2003 Apr 10;14(6):577-90. doi: 10.1089/104303403764539350. Hum Gene Ther. 2003. PMID: 12718767
-
Blockade of CD40 pathway enhances the induction of immune tolerance by immature dendritic cells genetically modified to express cytotoxic T lymphocyte antigen 4 immunoglobulin.Transplantation. 2003 Nov 15;76(9):1351-9. doi: 10.1097/01.TP.0000083557.25887.EE. Transplantation. 2003. PMID: 14627915
-
Tracking Donor-Reactive T Cells: Perspectives for the Development of Tolerance Protocols.Transplantation. 2015 Dec;99(12):2436-7. doi: 10.1097/TP.0000000000000999. Transplantation. 2015. PMID: 26627670 Review. No abstract available.
Cited by
-
Contributions of direct and indirect alloresponses to chronic rejection of kidney allografts in nonhuman primates.J Immunol. 2011 Nov 1;187(9):4589-97. doi: 10.4049/jimmunol.1003253. Epub 2011 Sep 28. J Immunol. 2011. PMID: 21957140 Free PMC article.
-
Engineered regulatory T cells coexpressing MHC class II:peptide complexes are efficient inhibitors of autoimmune T cell function and prevent the development of autoimmune arthritis.J Immunol. 2013 Jun 1;190(11):5382-91. doi: 10.4049/jimmunol.1300024. Epub 2013 Apr 29. J Immunol. 2013. PMID: 23630354 Free PMC article.
-
The importance of MHC class II in allogeneic bone marrow transplantation and chimerism-based solid organ tolerance in a rat model.PLoS One. 2020 May 22;15(5):e0233497. doi: 10.1371/journal.pone.0233497. eCollection 2020. PLoS One. 2020. PMID: 32442182 Free PMC article.
-
Safety and Efficacy of Allogeneic Lung Spheroid Cells in a Mismatched Rat Model of Pulmonary Fibrosis.Stem Cells Transl Med. 2017 Oct;6(10):1905-1916. doi: 10.1002/sctm.16-0374. Epub 2017 Aug 7. Stem Cells Transl Med. 2017. PMID: 28783251 Free PMC article.
-
Relevance of regulatory T cell promotion of donor-specific tolerance in solid organ transplantation.Front Immunol. 2012 Jul 13;3:184. doi: 10.3389/fimmu.2012.00184. eCollection 2012. Front Immunol. 2012. PMID: 22811678 Free PMC article.
References
-
- Starr TK, Jameson SC, Hogquist KA. Positive and negative selection of T cells. Annu Rev Immunol. 2003;21:139–176. - PubMed
-
- Sprent J, Kishimoto H. The thymus and negative selection. Immunol Rev. 2002;185:126–135. - PubMed
-
- Watanabe N, Wang YH, Lee HK, Ito T, Wang YH, Cao W, Liu YJ. Hassall’s corpuscles instruct dendritic cells to induce CD4+CD25+ regulatory T cells in human thymus. Nature. 2005;436:1181–1185. - PubMed
-
- Sakaguchi S. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat Immunol. 2005;6:345–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
