

RNAase-III enzyme Dicer maintains signaling pathways for differentiation and survival in mouse cortical neural stem cells
- PMID: 20103535
- PMCID: PMC2818196
- DOI: 10.1242/jcs.059659
RNAase-III enzyme Dicer maintains signaling pathways for differentiation and survival in mouse cortical neural stem cells
Abstract
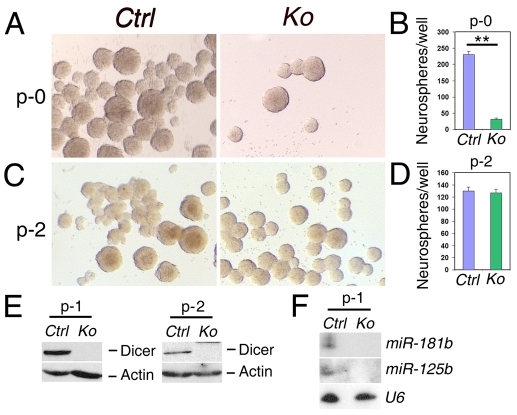
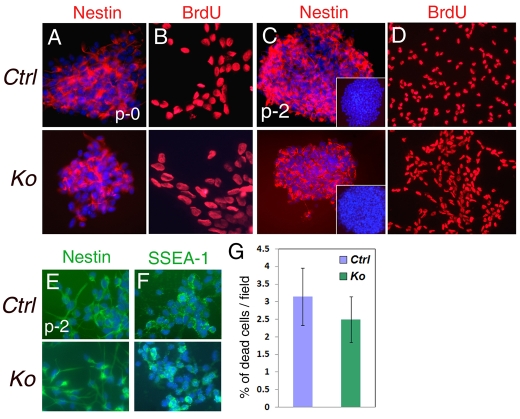
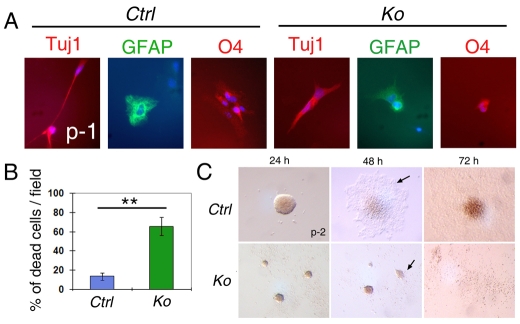
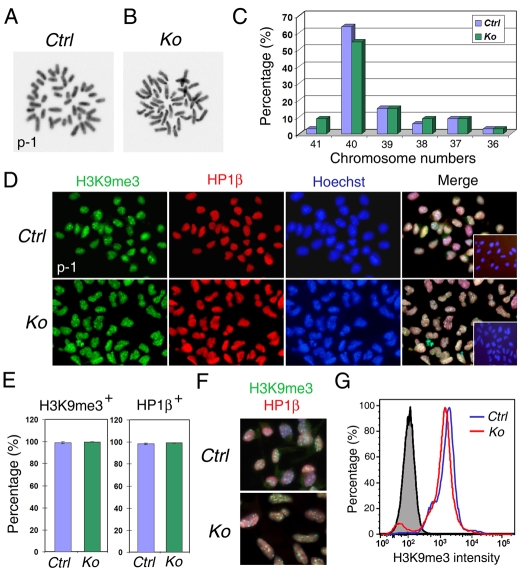
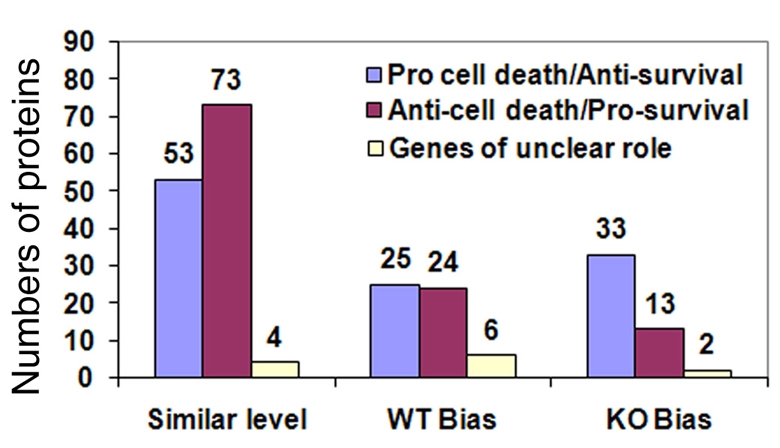
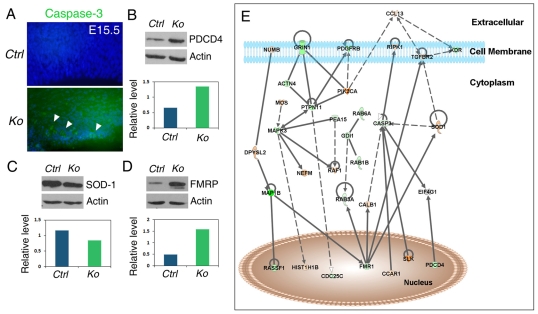
An important function of the RNAase-III enzyme Dicer is to process microRNA precursors into ~22-nucleotide non-coding small RNAs. But little is known about the role of Dicer in mammalian brain formation and neural stem cell (NSC) development. Here we show that Dicer plays a crucial role in controlling mouse cortical NSC development. We found that Dicer function is essential for expanding cortical neural progenitors and NSCs. We have identified a population of Dicer-deficient NSCs that can self-renew, and that display normal karyotype and heterochromatin protein expression levels but show enlarged nuclei. Dicer-deficient NSCs display abnormal differentiation and undergo cell death when mitogens are withdrawn. Dicer deletion affects the levels of many proteins, as revealed by a mass spectrometry proteomic approach. We have found that an increase of anti-survival and/or pro-apoptosis proteins and a decrease of pro-survival and/or anti-apoptosis proteins contribute to the cell death of Dicer-deficient NSCs, implying a general role for Dicer in protecting cells from apoptosis. Our results demonstrate important functions for Dicer in regulating NSC development by maintaining proper signaling pathways related to cell survival and differentiation.
Figures
Similar articles
-
Reversible block of mouse neural stem cell differentiation in the absence of dicer and microRNAs.PLoS One. 2010 Oct 18;5(10):e13453. doi: 10.1371/journal.pone.0013453. PLoS One. 2010. PMID: 20976144 Free PMC article.
-
Dicer is required for neural stem cell multipotency and lineage progression during cerebral cortex development.Neural Dev. 2013 Jul 29;8:14. doi: 10.1186/1749-8104-8-14. Neural Dev. 2013. PMID: 23895693 Free PMC article.
-
Dicer is required for proliferation, viability, migration and differentiation in corticoneurogenesis.Neuroscience. 2012 Oct 25;223:285-95. doi: 10.1016/j.neuroscience.2012.08.009. Epub 2012 Aug 13. Neuroscience. 2012. PMID: 22898830 Free PMC article.
-
Chromatin changes in dicer-deficient mouse embryonic stem cells in response to retinoic acid induced differentiation.PLoS One. 2013 Sep 9;8(9):e74556. doi: 10.1371/journal.pone.0074556. eCollection 2013. PLoS One. 2013. PMID: 24040281 Free PMC article.
-
Role of Dicer in female fertility.Trends Endocrinol Metab. 2009 Aug;20(6):265-72. doi: 10.1016/j.tem.2009.05.001. Epub 2009 Jul 29. Trends Endocrinol Metab. 2009. PMID: 19646895 Free PMC article. Review.
Cited by
-
Decoding the non-coding RNAs in Alzheimer's disease.Cell Mol Life Sci. 2012 Nov;69(21):3543-59. doi: 10.1007/s00018-012-1125-z. Epub 2012 Sep 6. Cell Mol Life Sci. 2012. PMID: 22955374 Free PMC article. Review.
-
MicroRNAs: regulators of neuronal fate.Curr Opin Cell Biol. 2013 Apr;25(2):215-21. doi: 10.1016/j.ceb.2012.12.007. Epub 2013 Jan 29. Curr Opin Cell Biol. 2013. PMID: 23374323 Free PMC article. Review.
-
The Zika Virus Capsid Disrupts Corticogenesis by Suppressing Dicer Activity and miRNA Biogenesis.Cell Stem Cell. 2020 Oct 1;27(4):618-632.e9. doi: 10.1016/j.stem.2020.07.012. Epub 2020 Aug 6. Cell Stem Cell. 2020. PMID: 32763144 Free PMC article.
-
Canonical microRNAs Enable Differentiation, Protect Against DNA Damage, and Promote Cholesterol Biosynthesis in Neural Stem Cells.Stem Cells Dev. 2017 Feb 1;26(3):177-188. doi: 10.1089/scd.2016.0259. Epub 2016 Dec 9. Stem Cells Dev. 2017. PMID: 27762676 Free PMC article.
-
Growth and folding of the mammalian cerebral cortex: from molecules to malformations.Nat Rev Neurosci. 2014 Apr;15(4):217-32. doi: 10.1038/nrn3707. Nat Rev Neurosci. 2014. PMID: 24646670 Free PMC article. Review.
References
-
- Anthony T. E., Klein C., Fishell G., Heintz N. (2004). Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron 41, 881-890 - PubMed
-
- Capela A., Temple S. (2006). LeX is expressed by principle progenitor cells in the embryonic nervous system, is secreted into their environment and binds Wnt-1. Dev. Biol. 291, 300-313 - PubMed
-
- Carmell M. A., Hannon G. J. (2004). RNase III enzymes and the initiation of gene silencing. Nat. Struct. Mol. Biol. 11, 214-218 - PubMed
-
- Chang T. C., Mendell J. T. (2007). microRNAs in vertebrate physiology and human disease. Annu. Rev. Genomics Hum. Genet. 8, 215-239 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
