

Repetitive behavior and increased activity in mice with Purkinje cell loss: a model for understanding the role of cerebellar pathology in autism
- PMID: 20105240
- PMCID: PMC2846709
- DOI: 10.1111/j.1460-9568.2009.07073.x
Repetitive behavior and increased activity in mice with Purkinje cell loss: a model for understanding the role of cerebellar pathology in autism
Abstract
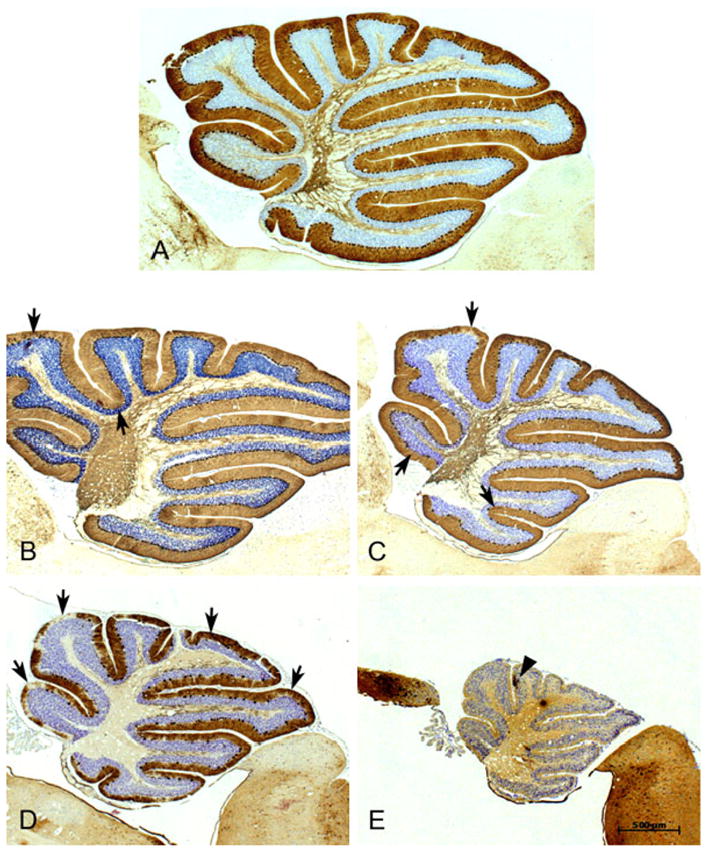
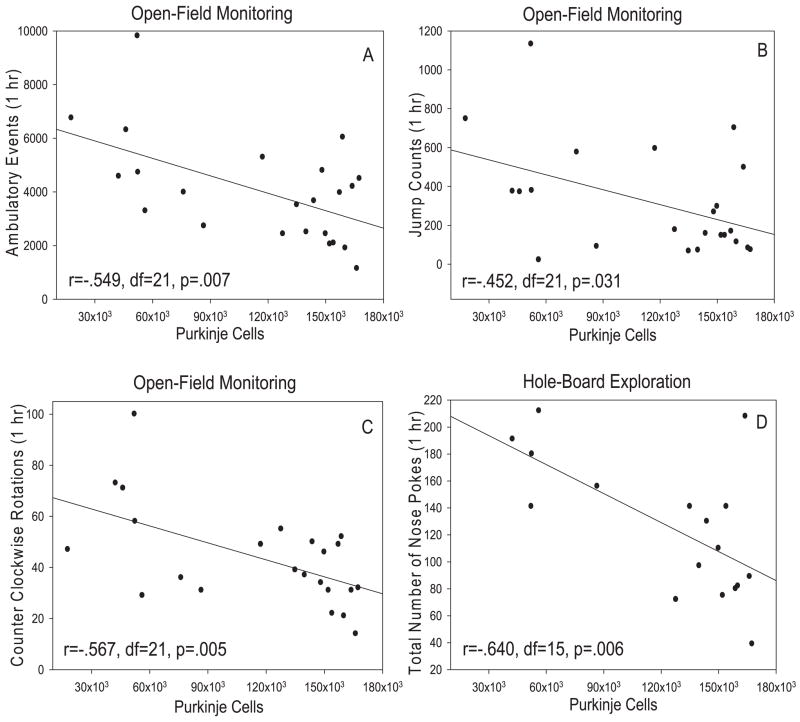
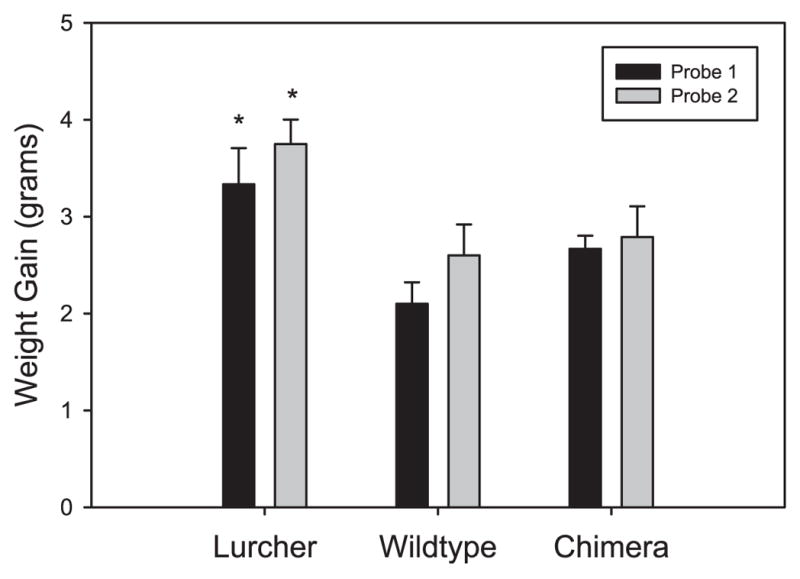
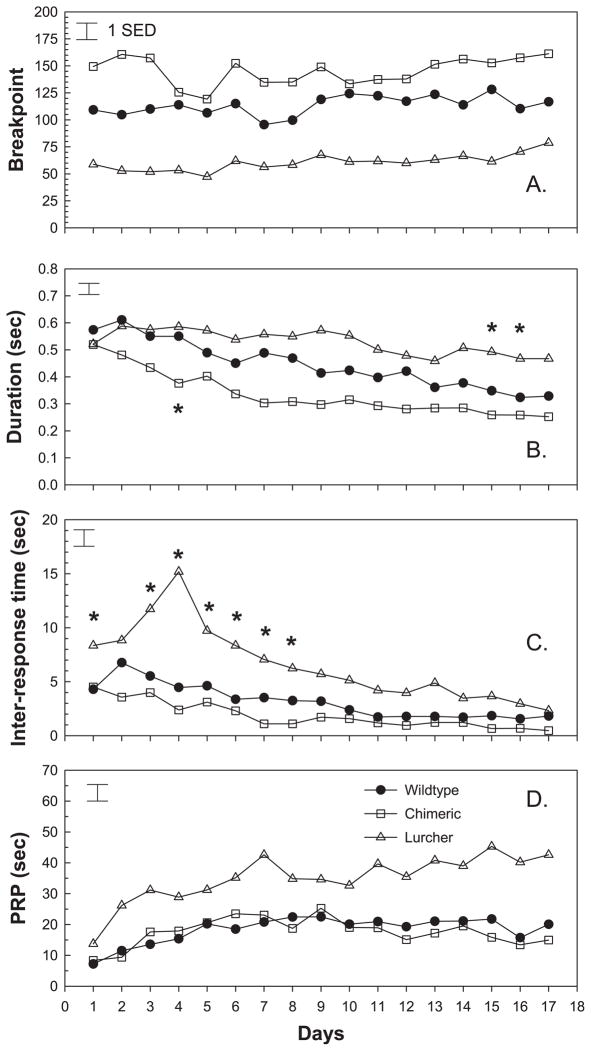
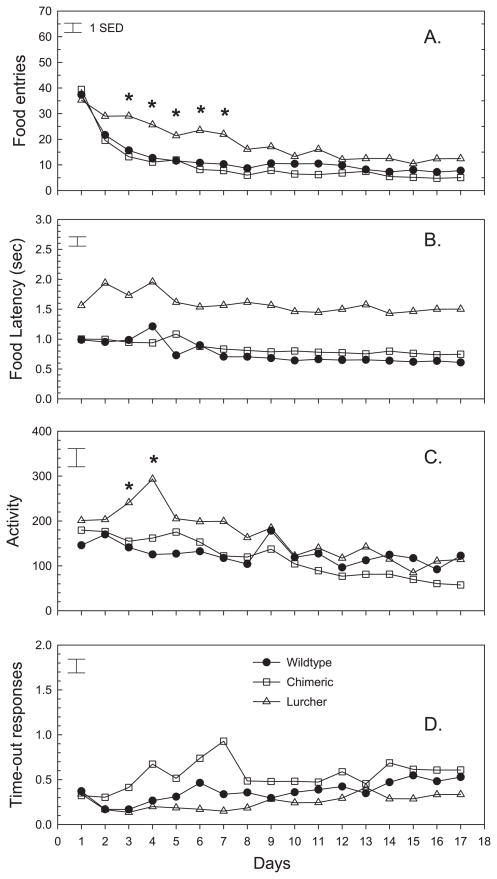
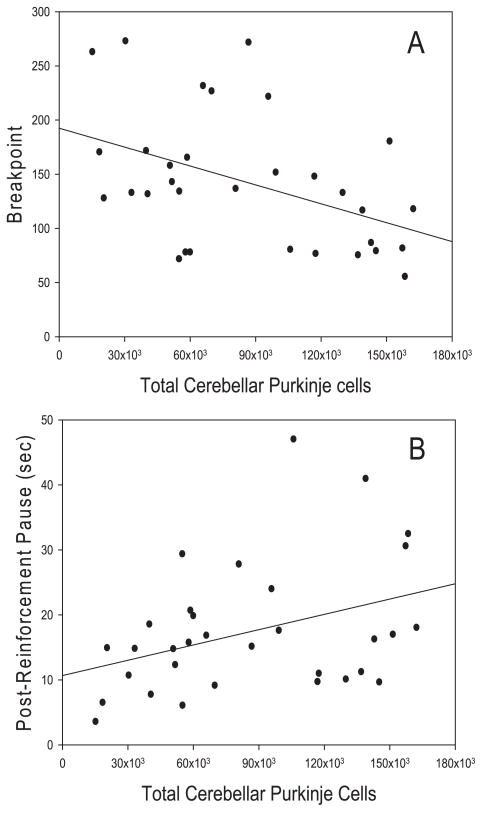
Repetitive behaviors and hyperactivity are common features of developmental disorders, including autism. Neuropathology of the cerebellum is also a frequent occurrence in autism and other developmental disorders. Recent studies have indicated that cerebellar pathology may play a causal role in the generation of repetitive and hyperactive behaviors. In this study, we examined the relationship between cerebellar pathology and these behaviors in a mouse model of Purkinje cell loss. Specifically, we made aggregation chimeras between Lc/+ mutant embryos and +/+ embryos. Lc/+ mice lose 100% of their Purkinje cells postnatally due to a cell-intrinsic gain-of-function mutation. Through our histological examination, we demonstrated that Lc/+<-->+/+ chimeric mice have Purkinje cells ranging from zero to normal numbers. Our analysis of these chimeric cerebella confirmed previous studies on Purkinje cell lineage. The results of both open-field activity and hole-board exploration testing indicated negative relationships between Purkinje cell number and measures of activity and investigatory nose-poking. Additionally, in a progressive-ratio operant paradigm, we found that Lc/+ mice lever-pressed significantly less than +/+ controls, which led to significantly lower breakpoints in this group. In contrast, chimeric mice lever-pressed significantly more than controls and this repetitive lever-pressing behavior was significantly and negatively correlated with total Purkinje cell numbers. Although the performance of Lc/+ mice is probably related to their motor deficits, the significant relationships between Purkinje cell number and repetitive lever-pressing behavior as well as open-field activity measures provide support for a role of cerebellar pathology in generating repetitive behavior and increased activity in chimeric mice.
Figures
References
-
- Abercrombie M. Estimation of nuclear population from microtome sections. Anat Rec. 1946;94:239–247. - PubMed
-
- Baader SL, Schilling ML, Rosengarten B, Pretsch W, Teutsch HF, Oberdick J, Schilling K. Purkinje cell lineage and the topographic organization of the cerebellar cortex: a view from X inactivation mosaics. Dev Biol. 1996;174:393–406. - PubMed
-
- Bleuler EP. Dementia Praecox or the Group of Schizophrenias. International University Press; New York: 1950.
-
- Bodfish JW, Symons FJ, Parker DE, Lewis MH. Varieties of repetitive behavior in autism: comparisons to mental retardation. J Autism Dev Disord. 2000;30:237–243. - PubMed
-
- Boltshauser E, Steinlin M, Martin E, Deonna T. Unilateral cerebellar aplasia. Neuropediatrics. 1996;27:50–53. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
