

Basal ganglia control of sleep-wake behavior and cortical activation
- PMID: 20105243
- PMCID: PMC3928571
- DOI: 10.1111/j.1460-9568.2009.07062.x
Basal ganglia control of sleep-wake behavior and cortical activation
Abstract
The basal ganglia (BG) are involved in numerous neurobiological processes that operate on the basis of wakefulness, including motor function, learning, emotion and addictive behaviors. We hypothesized that the BG might play an important role in the regulation of wakefulness. To test this prediction, we made cell body-specific lesions in the striatum and globus pallidus (GP) using ibotenic acid. We found that rats with striatal (caudoputamen) lesions exhibited a 14.95% reduction in wakefulness and robust fragmentation of sleep-wake behavior, i.e. an increased number of state transitions and loss of ultra-long wake bouts (> 120 min). These lesions also resulted in a reduction in the diurnal variation of sleep-wakefulness. On the other hand, lesions of the accumbens core resulted in a 26.72% increase in wakefulness and a reduction in non-rapid eye movement (NREM) sleep bout duration. In addition, rats with accumbens core lesions exhibited excessive digging and scratching. GP lesions also produced a robust increase in wakefulness (45.52%), and frequent sleep-wake transitions and a concomitant decrease in NREM sleep bout duration. Lesions of the subthalamic nucleus or the substantia nigra reticular nucleus produced only minor changes in the amount of sleep-wakefulness and did not alter sleep architecture. Finally, power spectral analysis revealed that lesions of the striatum, accumbens and GP slowed down the cortical electroencephalogram. Collectively, our results suggest that the BG, via a cortico-striato-pallidal loop, are important neural circuitry regulating sleep-wake behaviors and cortical activation.
Figures



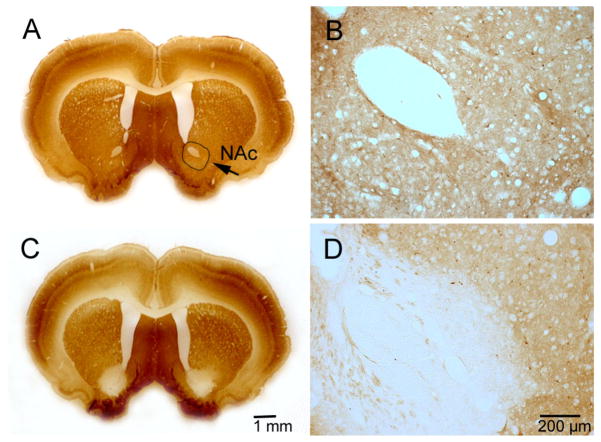


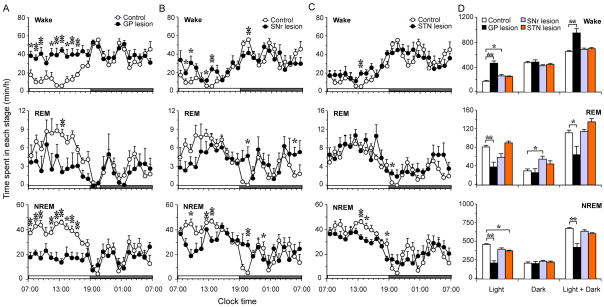

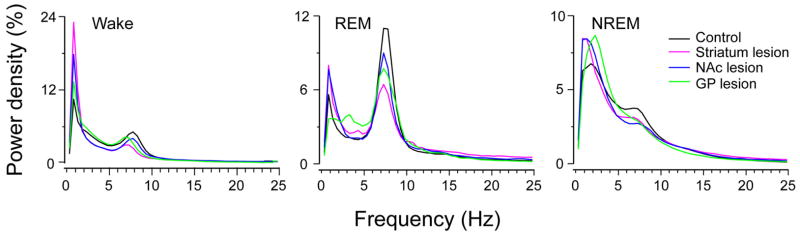

Similar articles
-
Lesion of the pedunculopontine tegmental nucleus in rat augments cortical activation and disturbs sleep/wake state transitions structure.Exp Neurol. 2013 Sep;247:562-71. doi: 10.1016/j.expneurol.2013.02.007. Epub 2013 Feb 26. Exp Neurol. 2013. PMID: 23481548
-
Nigrostriatal Dopamine Acting on Globus Pallidus Regulates Sleep.Cereb Cortex. 2016 Apr;26(4):1430-9. doi: 10.1093/cercor/bhu241. Epub 2014 Oct 14. Cereb Cortex. 2016. PMID: 25316334 Free PMC article.
-
Unrelated course of subthalamic nucleus and globus pallidus neuronal activities across vigilance states in the rat.Eur J Neurosci. 2000 Sep;12(9):3361-74. doi: 10.1046/j.1460-9568.2000.00199.x. Eur J Neurosci. 2000. PMID: 10998119
-
Role of the basal ganglia in the control of sleep and wakefulness.Curr Opin Neurobiol. 2013 Oct;23(5):780-5. doi: 10.1016/j.conb.2013.02.001. Epub 2013 Apr 24. Curr Opin Neurobiol. 2013. PMID: 23465424 Free PMC article. Review.
-
[Selective stimulations and lesions of the rat brain nuclei as the models for research of the human sleep pathology mechanisms].Glas Srp Akad Nauka Med. 2011;(51):85-97. Glas Srp Akad Nauka Med. 2011. PMID: 22165729 Review. Serbian.
Cited by
-
Perspectives on the rapid eye movement sleep switch in rapid eye movement sleep behavior disorder.Sleep Med. 2013 Aug;14(8):707-13. doi: 10.1016/j.sleep.2013.03.017. Epub 2013 Jun 13. Sleep Med. 2013. PMID: 23768838 Free PMC article. Review.
-
Control of sleep and wakefulness.Physiol Rev. 2012 Jul;92(3):1087-187. doi: 10.1152/physrev.00032.2011. Physiol Rev. 2012. PMID: 22811426 Free PMC article. Review.
-
Optogenetic Stimulation of GABAergic Neurons in the Globus Pallidus Produces Hyperkinesia.Front Behav Neurosci. 2018 Aug 27;12:185. doi: 10.3389/fnbeh.2018.00185. eCollection 2018. Front Behav Neurosci. 2018. PMID: 30210317 Free PMC article.
-
Deep structural brain lesions associated with consciousness impairment early after hemorrhagic stroke.Sci Rep. 2019 Mar 12;9(1):4174. doi: 10.1038/s41598-019-41042-2. Sci Rep. 2019. PMID: 30862910 Free PMC article.
-
Early-life stress alters sleep structure and the excitatory-inhibitory balance in the nucleus accumbens in aged mice.Chin Med J (Engl). 2019 Jul 5;132(13):1582-1590. doi: 10.1097/CM9.0000000000000279. Chin Med J (Engl). 2019. PMID: 31045908 Free PMC article.
References
-
- Gerashchenko D, Blanco-Centurion C, Greco MA, Shiromani PJ. Effects of lateral hypothalamic lesion with the neurotoxin hypocretin-2-saporin on sleep in Long-Evans rats. Neuroscience. 2003;116:223–235. - PubMed
-
- Gerashchenko D, Blanco-Centurion CA, Miller JD, Shiromani PJ. Insomnia following hypocretin2-saporin lesions of the substantia nigra. Neuroscience. 2006;137:29–36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
