

Sulfotyrosines of the Kaposi's sarcoma-associated herpesvirus G protein-coupled receptor promote tumorigenesis through autocrine activation
- PMID: 20106924
- PMCID: PMC2838108
- DOI: 10.1128/JVI.01939-09
Sulfotyrosines of the Kaposi's sarcoma-associated herpesvirus G protein-coupled receptor promote tumorigenesis through autocrine activation
Abstract
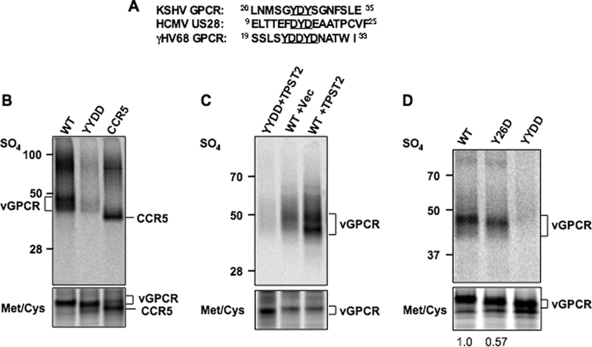
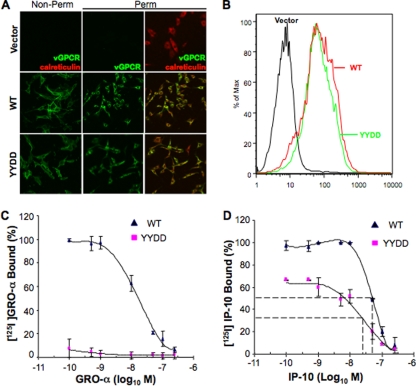
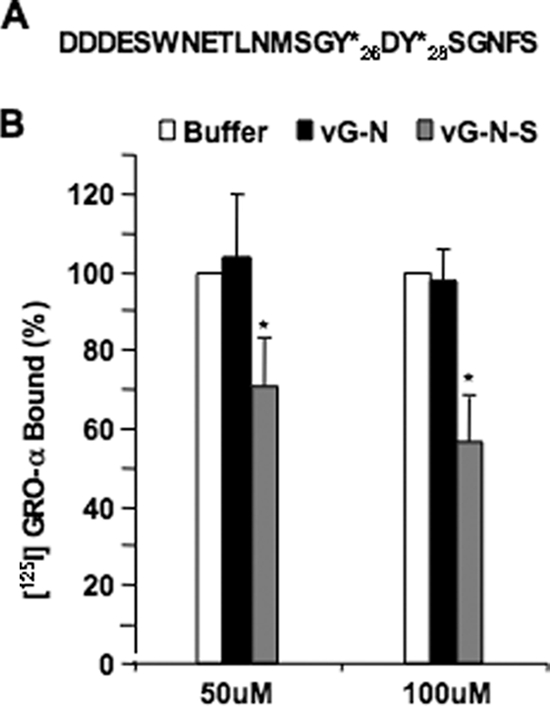
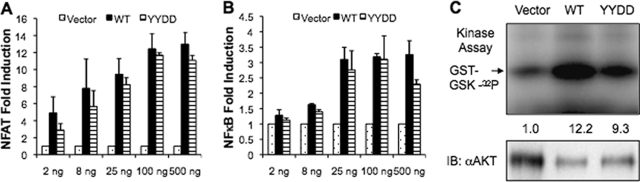
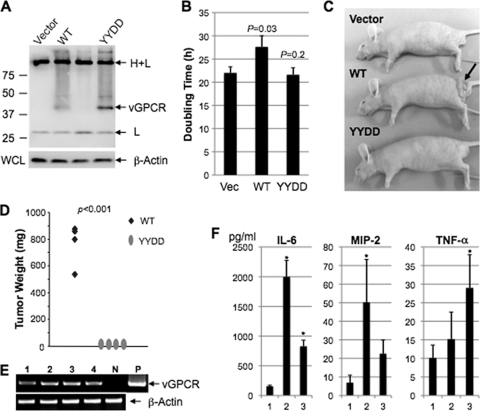
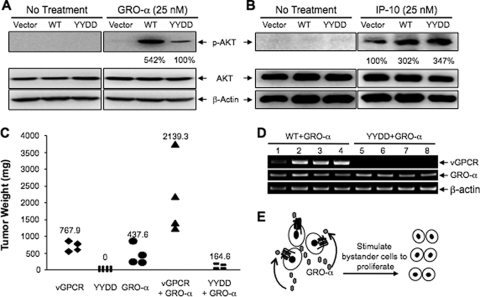
The Kaposi's sarcoma-associated herpesvirus (KSHV) G protein-coupled receptor (vGPCR) is a bona fide signaling molecule that is implicated in KSHV-associated malignancies. Whereas vGPCR activates specific cellular signaling pathways in a chemokine-independent fashion, vGPCR binds a broad spectrum of CC and CXC chemokines, and the roles of chemokines in vGPCR tumorigenesis remain poorly understood. We report here that vGPCR is posttranslationally modified by sulfate groups at tyrosine residues within its N-terminal extracellular domain. A chemokine-binding assay demonstrated that the tyrosine sulfate moieties were critical for vGPCR association with GRO-alpha (an agonist) but not with IP-10 (an inverse agonist). A sulfated peptide corresponding to residues 12 through 33 of vGPCR, but not the unsulfated equivalent, partially inhibited vGPCR association with GRO-alpha. Although the vGPCR variant lacking sulfotyrosines activated downstream signaling pathways, the ability of the unsulfated vGPCR variant to induce tumor growth in nude mice was significantly diminished. Furthermore, the unsulfated vGPCR variant was unable to induce the secretion of proliferative cytokines, some of which serve as vGPCR agonists. This implies that autocrine activation by agonist chemokines is critical for vGPCR tumorigenesis. Indeed, GRO-alpha increased vGPCR-mediated AKT phosphorylation and vGPCR tumorigenesis in a sulfotyrosine-dependent manner. Our findings support the conclusion that autocrine activation triggered by chemokine agonists via sulfotyrosines is necessary for vGPCR tumorigenesis, thereby providing a rationale for future therapeutic design targeting the tumorigenic vGPCR.
Figures
Similar articles
-
The unsulfated extracellular N-terminus of vGPCR reduces the tumorigenicity of hGRO-α in nude mice.Sci China Life Sci. 2013 Jan;56(1):26-31. doi: 10.1007/s11427-012-4405-9. Epub 2012 Dec 27. Sci China Life Sci. 2013. PMID: 23269549
-
KSHV G-protein coupled receptor vGPCR oncogenic signaling upregulation of Cyclooxygenase-2 expression mediates angiogenesis and tumorigenesis in Kaposi's sarcoma.PLoS Pathog. 2020 Oct 15;16(10):e1009006. doi: 10.1371/journal.ppat.1009006. eCollection 2020 Oct. PLoS Pathog. 2020. PMID: 33057440 Free PMC article.
-
The Kaposi's sarcoma-associated herpesvirus G protein-coupled receptor as a therapeutic target for the treatment of Kaposi's sarcoma.Cancer Res. 2006 Jan 1;66(1):168-74. doi: 10.1158/0008-5472.CAN-05-1026. Cancer Res. 2006. PMID: 16397229
-
KSHV non-structural membrane proteins involved in the activation of intracellular signaling pathways and the pathogenesis of Kaposi's sarcoma.Curr Opin Virol. 2016 Oct;20:11-19. doi: 10.1016/j.coviro.2016.07.008. Epub 2016 Aug 9. Curr Opin Virol. 2016. PMID: 27518127 Review.
-
Akt/TSC/mTOR activation by the KSHV G protein-coupled receptor: emerging insights into the molecular oncogenesis and treatment of Kaposi's sarcoma.Cell Cycle. 2007 Feb 15;6(4):438-43. doi: 10.4161/cc.6.4.3843. Epub 2007 Feb 12. Cell Cycle. 2007. PMID: 17329974 Review.
Cited by
-
Glycosylation of KSHV encoded vGPCR functions in its signaling and tumorigenicity.Viruses. 2015 Mar 31;7(4):1627-41. doi: 10.3390/v7041627. Viruses. 2015. PMID: 25835533 Free PMC article.
-
In vivo electroporation and non-protein based screening assays to identify antibodies against native protein conformations.Hybridoma (Larchmt). 2011 Oct;30(5):409-18. doi: 10.1089/hyb.2010.0120. Hybridoma (Larchmt). 2011. PMID: 22008067 Free PMC article.
-
The Viral G Protein-Coupled Receptor ORF74 Hijacks β-Arrestins for Endocytic Trafficking in Response to Human Chemokines.PLoS One. 2015 Apr 20;10(4):e0124486. doi: 10.1371/journal.pone.0124486. eCollection 2015. PLoS One. 2015. PMID: 25894435 Free PMC article.
-
Modulation of cellular signaling by herpesvirus-encoded G protein-coupled receptors.Front Pharmacol. 2015 Mar 9;6:40. doi: 10.3389/fphar.2015.00040. eCollection 2015. Front Pharmacol. 2015. PMID: 25805993 Free PMC article. Review.
-
Molecular mechanisms deployed by virally encoded G protein-coupled receptors in human diseases.Annu Rev Pharmacol Toxicol. 2013;53:331-54. doi: 10.1146/annurev-pharmtox-010510-100608. Epub 2012 Oct 22. Annu Rev Pharmacol Toxicol. 2013. PMID: 23092247 Free PMC article. Review.
References
-
- Reference deleted.
-
- Arvanitakis, L., E. Geras-Raaka, A. Varma, M. C. Gershengorn, and E. Cesarman. 1997. Human herpesvirus KSHV encodes a constitutively active G-protein-coupled receptor linked to cell proliferation. Nature 385:347-350. - PubMed
-
- Bais, C., B. Santomasso, O. Coso, L. Arvanitakis, E. G. Raaka, J. S. Gutkind, A. S. Asch, E. Cesarman, M. C. Gershengorn, and E. A. Mesri. 1998. G-protein-coupled receptor of Kaposi's sarcoma-associated herpesvirus is a viral oncogene and angiogenesis activator. Nature 391:86-89. - PubMed
-
- Bais, C., A. Van Geelen, P. Eroles, A. Mutlu, C. Chiozzini, S. Dias, R. L. Silverstein, S. Rafii, and E. A. Mesri. 2003. Kaposi's sarcoma associated herpesvirus G protein-coupled receptor immortalizes human endothelial cells by activation of the VEGF receptor-2/ KDR. Cancer Cell 3:131-143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
