

Trypanosoma cruzi subverts host cell sialylation and may compromise antigen-specific CD8+ T cell responses
- PMID: 20106975
- PMCID: PMC2859498
- DOI: 10.1074/jbc.M109.096305
Trypanosoma cruzi subverts host cell sialylation and may compromise antigen-specific CD8+ T cell responses
Abstract
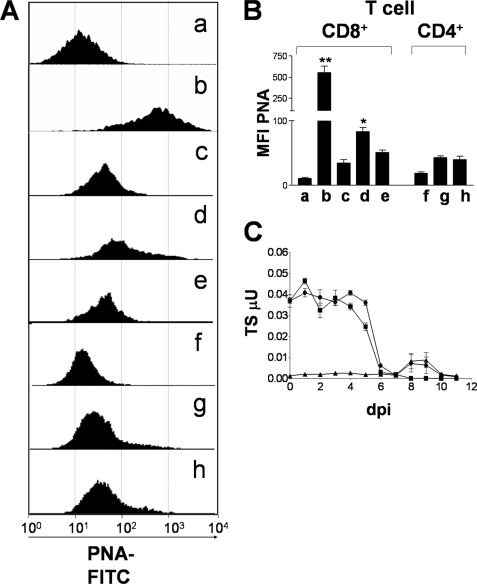
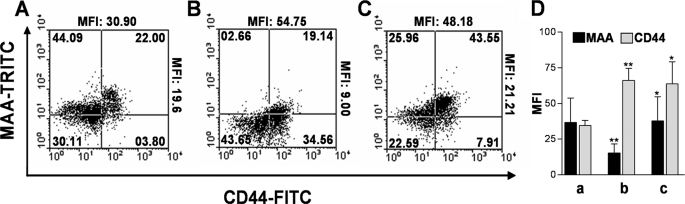
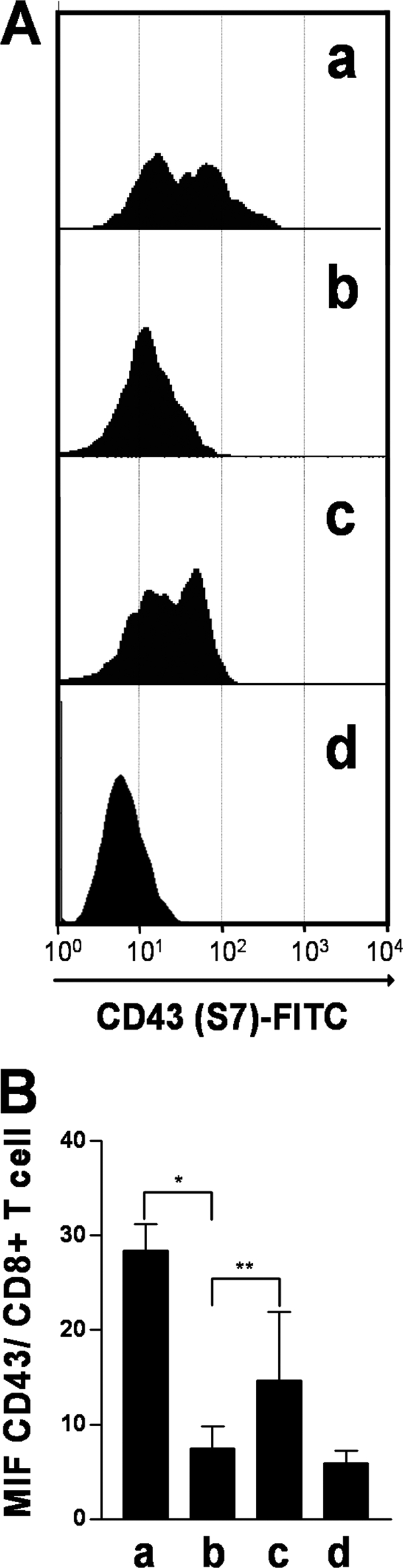
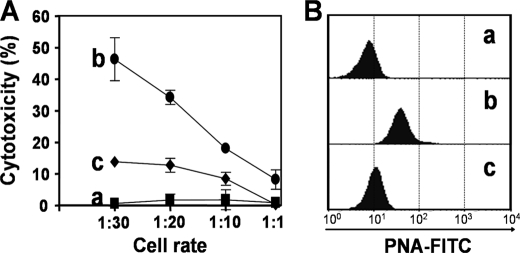
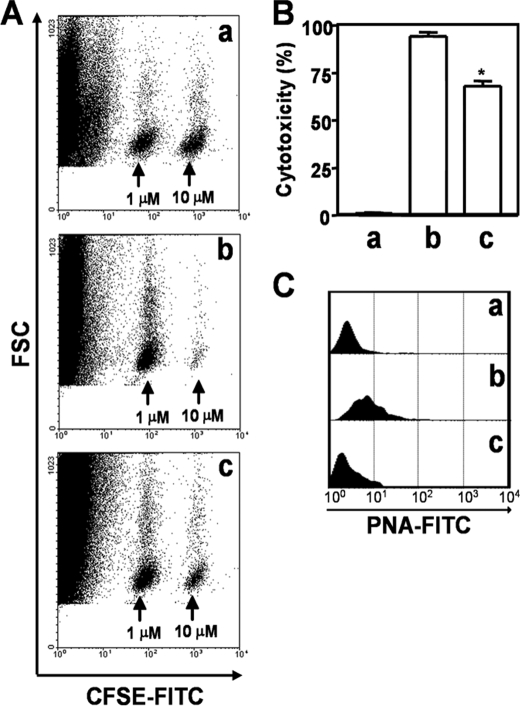
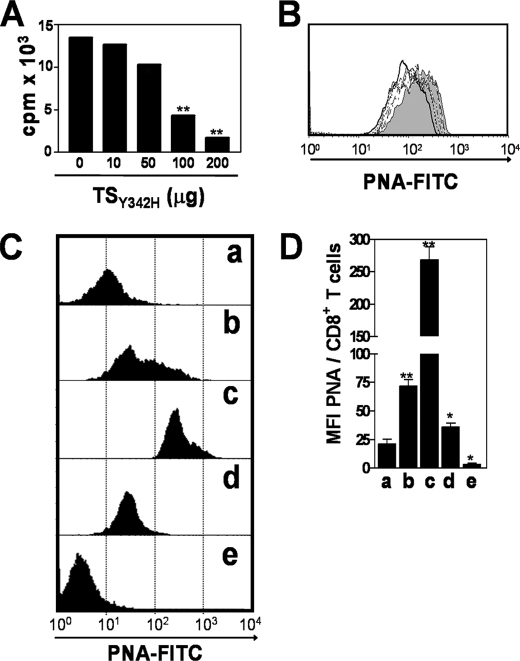
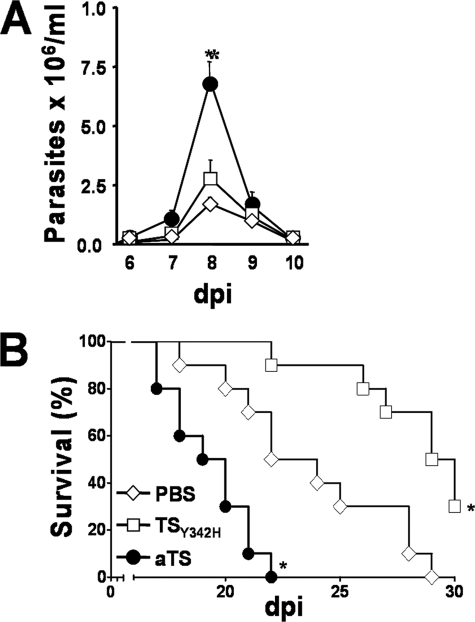
Upon activation, cytotoxic CD8(+) T lymphocytes are desialylated exposing beta-galactose residues in a physiological change that enhances their effector activity and that can be monitored on the basis of increased binding of the lectin peanut agglutinin. Herein, we investigated the impact of sialylation mediated by trans-sialidase, a specific and unique Trypanosoma transglycosylase for sialic acid, on CD8(+) T cell response of mice infected with T. cruzi. Our data demonstrate that T. cruzi uses its trans-sialidase enzyme to resialylate the CD8(+) T cell surface, thereby dampening antigen-specific CD8(+) T cell response that might favor its own persistence in the mammalian host. Binding of the monoclonal antibody S7, which recognizes sialic acid-containing epitopes on the 115-kDa isoform of CD43, was augmented on CD8(+) T cells from ST3Gal-I-deficient infected mice, indicating that CD43 is one sialic acid acceptor for trans-sialidase activity on the CD8(+) T cell surface. The cytotoxic activity of antigen-experienced CD8(+) T cells against the immunodominant trans-sialidase synthetic peptide IYNVGQVSI was decreased following active trans-sialidase-mediated resialylation in vitro and in vivo. Inhibition of the parasite's native trans-sialidase activity during infection strongly decreased CD8(+) T cell sialylation, reverting it to the glycosylation status expected in the absence of parasite manipulation increasing mouse survival. Taken together, these results demonstrate, for the first time, that T. cruzi subverts sialylation to attenuate CD8(+) T cell interactions with peptide-major histocompatibility complex class I complexes. CD8(+) T cell resialylation may represent a sophisticated strategy to ensure lifetime host parasitism.
Figures
Similar articles
-
Trans-sialidase recombinant protein mixed with CpG motif-containing oligodeoxynucleotide induces protective mucosal and systemic trypanosoma cruzi immunity involving CD8+ CTL and B cell-mediated cross-priming.J Immunol. 2007 Nov 15;179(10):6889-900. doi: 10.4049/jimmunol.179.10.6889. J Immunol. 2007. PMID: 17982080
-
Enzymatically inactive trans-sialidase from Trypanosoma cruzi binds sialyl and beta-galactopyranosyl residues in a sequential ordered mechanism.J Biol Chem. 2004 Feb 13;279(7):5323-8. doi: 10.1074/jbc.M310663200. Epub 2003 Nov 21. J Biol Chem. 2004. PMID: 14634017
-
Costimulatory Effects of an Immunodominant Parasite Antigen Paradoxically Prevent Induction of Optimal CD8 T Cell Protective Immunity.PLoS Pathog. 2016 Sep 19;12(9):e1005896. doi: 10.1371/journal.ppat.1005896. eCollection 2016 Sep. PLoS Pathog. 2016. PMID: 27642757 Free PMC article.
-
Theft and Reception of Host Cell's Sialic Acid: Dynamics of Trypanosoma Cruzi Trans-sialidases and Mucin-Like Molecules on Chagas' Disease Immunomodulation.Front Immunol. 2019 Feb 6;10:164. doi: 10.3389/fimmu.2019.00164. eCollection 2019. Front Immunol. 2019. PMID: 30787935 Free PMC article. Review.
-
Trans-sialidase, SAPA amino acid repeats and the relationship between Trypanosoma cruzi and the mammalian host.Parasitology. 1994;108 Suppl:S37-44. doi: 10.1017/s0031182000075703. Parasitology. 1994. PMID: 8084653 Review.
Cited by
-
Subdominant/cryptic CD8 T cell epitopes contribute to resistance against experimental infection with a human protozoan parasite.PLoS One. 2011;6(7):e22011. doi: 10.1371/journal.pone.0022011. Epub 2011 Jul 14. PLoS One. 2011. PMID: 21779365 Free PMC article.
-
Glycomics in Human Diseases and Its Emerging Role in Biomarker Discovery.Biomedicines. 2025 Aug 21;13(8):2034. doi: 10.3390/biomedicines13082034. Biomedicines. 2025. PMID: 40868286 Free PMC article. Review.
-
Overview of the role of kinetoplastid surface carbohydrates in infection and host cell invasion: prospects for therapeutic intervention.Parasitology. 2019 Dec;146(14):1743-1754. doi: 10.1017/S0031182019001355. Epub 2019 Oct 11. Parasitology. 2019. PMID: 31603063 Free PMC article. Review.
-
ST8Sia2 polysialyltransferase protects against infection by Trypanosoma cruzi.PLoS Negl Trop Dis. 2024 Sep 25;18(9):e0012454. doi: 10.1371/journal.pntd.0012454. eCollection 2024 Sep. PLoS Negl Trop Dis. 2024. PMID: 39321148 Free PMC article.
-
Evidence of ternary complex formation in Trypanosoma cruzi trans-sialidase catalysis.J Biol Chem. 2014 Jan 3;289(1):423-36. doi: 10.1074/jbc.M112.399303. Epub 2013 Nov 5. J Biol Chem. 2014. PMID: 24194520 Free PMC article.
References
-
- Moody A. M., Chui D., Reche P. A., Priatel J. J., Marth J. D., Reinherz E. L. (2001) Cell 107, 501–512 - PubMed
-
- Daniels M. A., Devine L., Miller J. D., Moser J. M., Lukacher A. E., Altman J. D., Kavathas P., Hogquist K. A., Jameson S. C. (2001) Immunity 15, 1051–1061 - PubMed
-
- Moody A. M., North S. J., Reinhold B., Van Dyken S. J., Rogers M. E., Panico M., Dell A., Morris H. R., Marth J. D., Reinherz E. L. (2003) J. Biol. Chem. 278, 7240–7246 - PubMed
-
- Galvan M., Murali-Krishna K., Ming L. L., Baum L., Ahmed R. (1998) J. Immunol. 161, 641–648 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
