

Retinophilin is a light-regulated phosphoprotein required to suppress photoreceptor dark noise in Drosophila
- PMID: 20107052
- PMCID: PMC2847805
- DOI: 10.1523/JNEUROSCI.4464-09.2010
Retinophilin is a light-regulated phosphoprotein required to suppress photoreceptor dark noise in Drosophila
Abstract
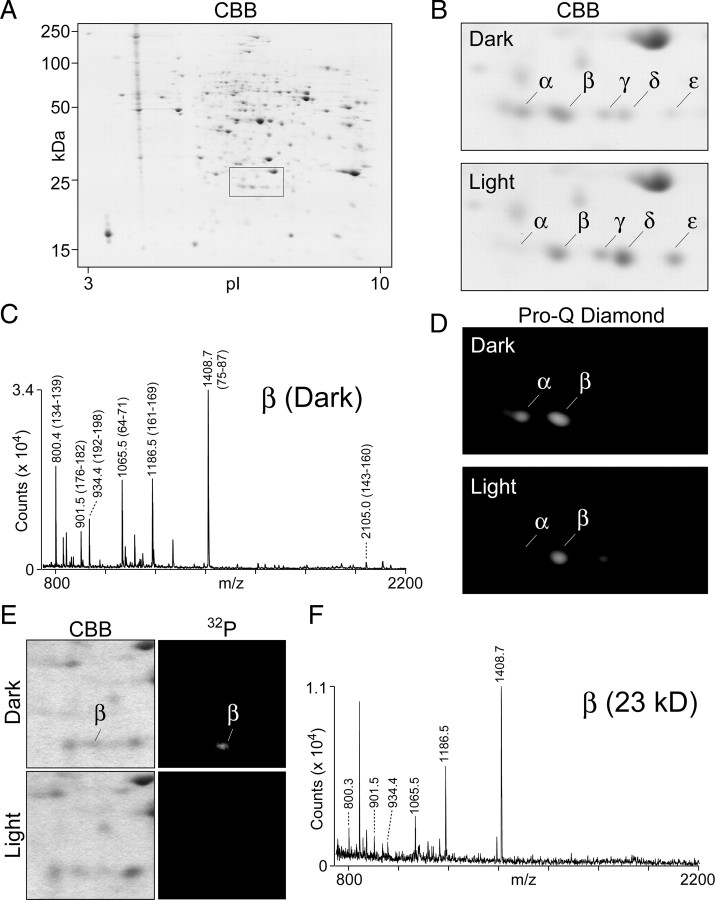
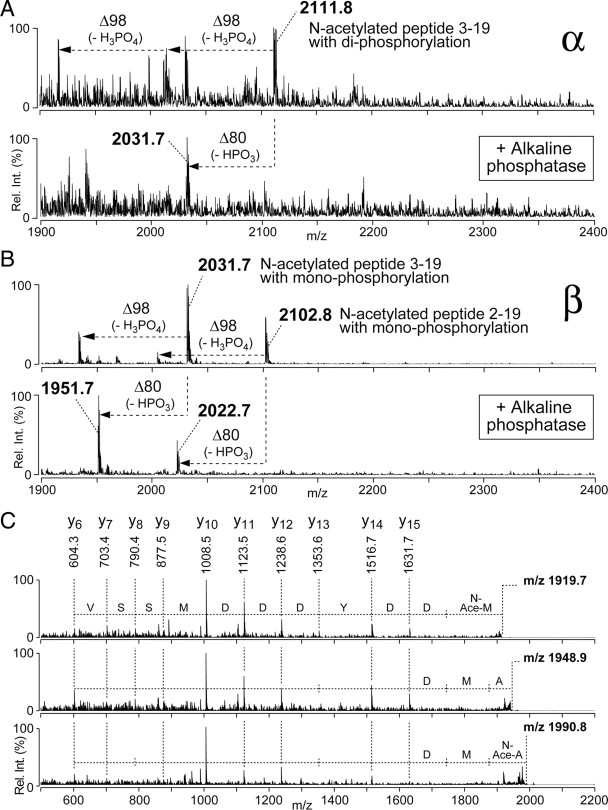
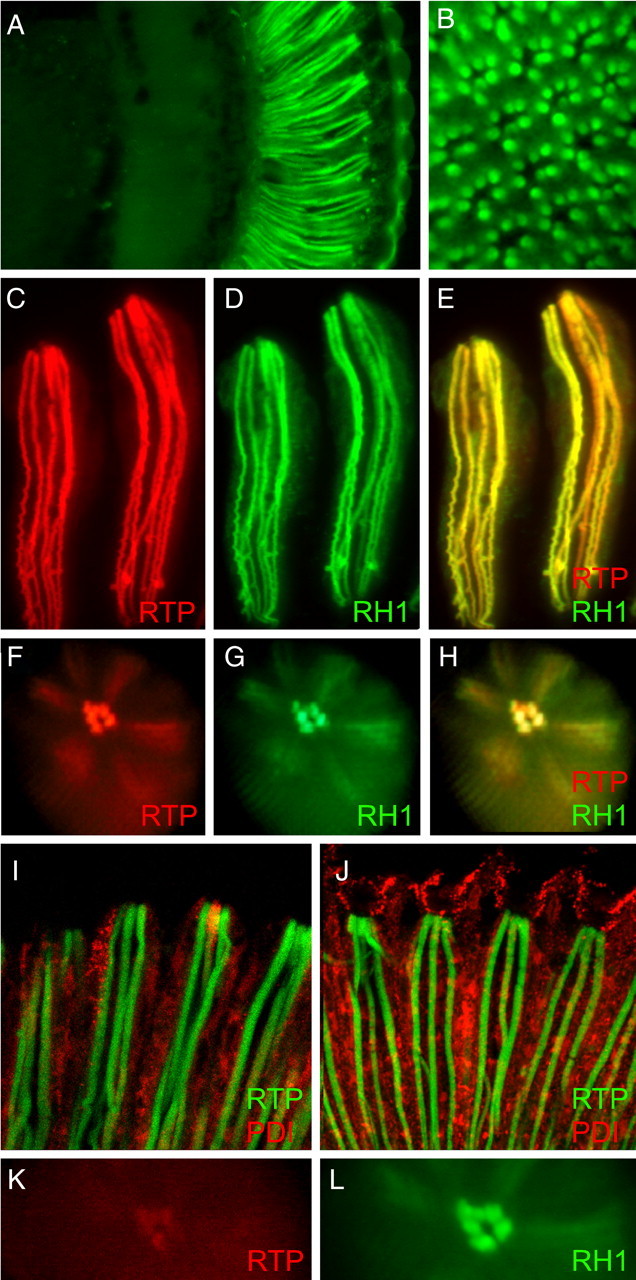
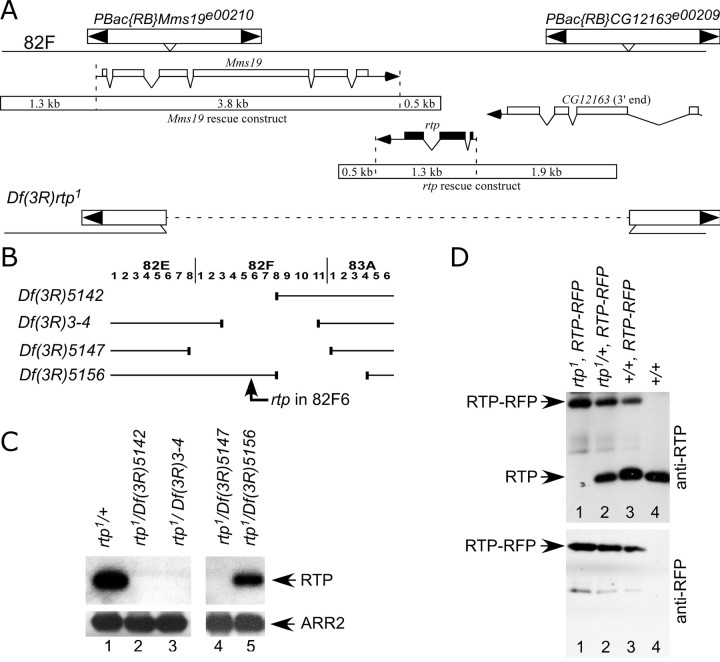
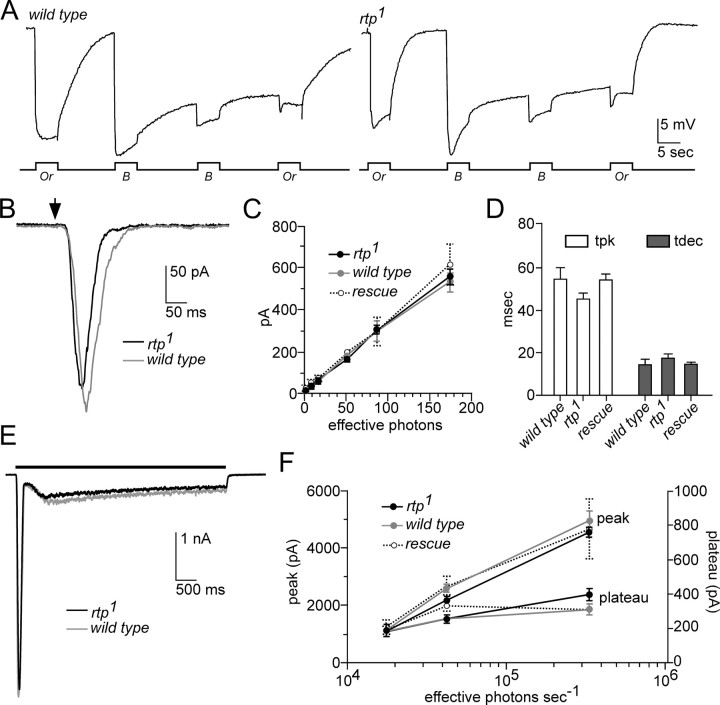
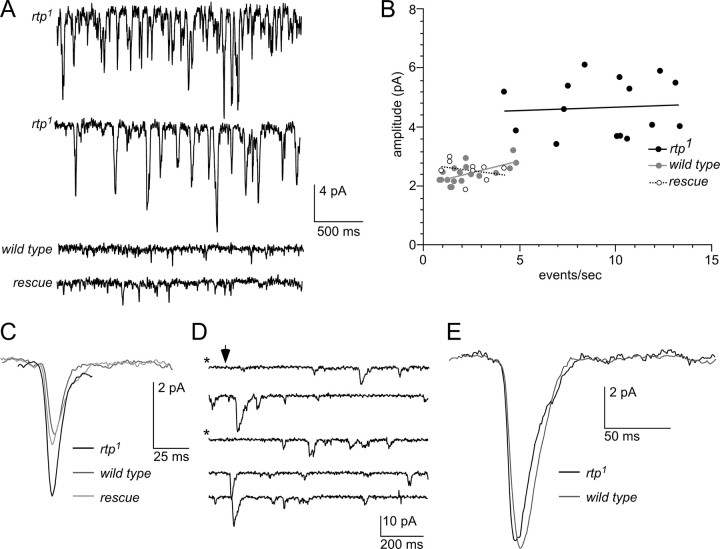
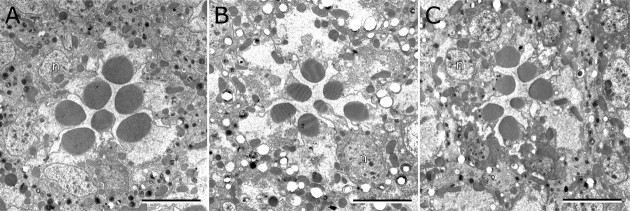
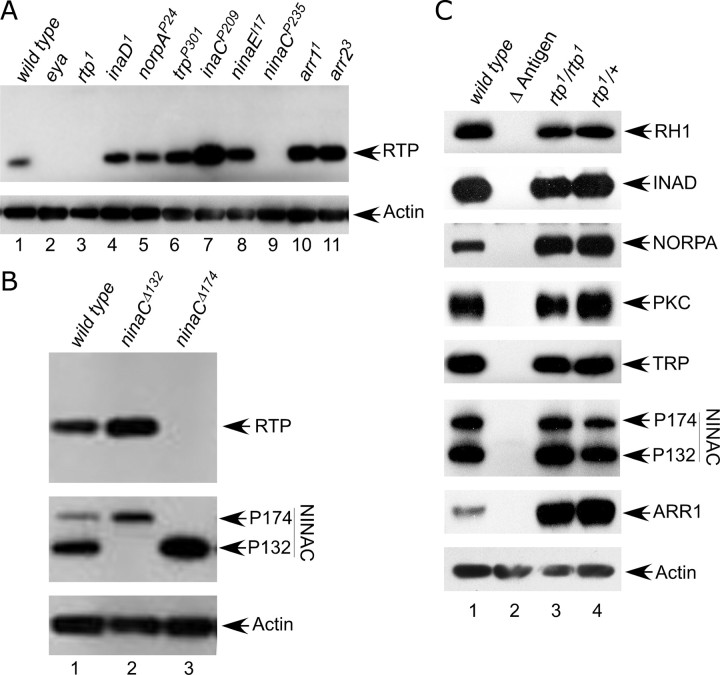
Photoreceptor cells achieve high sensitivity, reliably detecting single photons, while limiting the spontaneous activation events responsible for dark noise. We used proteomic, genetic, and electrophysiological approaches to characterize Retinophilin (RTP) (CG10233) in Drosophila photoreceptors and establish its involvement in dark-noise suppression. RTP possesses membrane occupation and recognition nexus (MORN) motifs, a structure shared with mammalian junctophilins and other membrane-associated proteins found within excitable cells. We show the MORN repeats, and both the N- and C-terminal domains, are required for RTP localization in the microvillar light-gathering organelle, the rhabdomere. RTP exists in multiple phosphorylated isoforms under dark conditions and is dephosphorylated by light exposure. An RTP deletion mutant exhibits a high rate of spontaneous membrane depolarization events in dark conditions but retains the normal kinetics of the light response. Photoreceptors lacking neither inactivation nor afterpotential C (NINAC) myosin III, a motor protein/kinase, also display a similar dark-noise phenotype as the RTP deletion. We show that NINAC mutants are depleted for RTP. These results suggest the increase in dark noise in NINAC mutants is attributable to lack of RTP and, furthermore, defines a novel role for NINAC in the rhabdomere. We propose that RTP is a light-regulated phosphoprotein that organizes rhabdomeric components to suppress random activation of the phototransduction cascade and thus increases the signaling fidelity of dark-adapted photoreceptors.
Figures
Similar articles
-
PDA (prolonged depolarizing afterpotential)-defective mutants: the story of nina's and ina's--pinta and santa maria, too.J Neurogenet. 2012 Jun;26(2):216-37. doi: 10.3109/01677063.2011.642430. Epub 2012 Jan 27. J Neurogenet. 2012. PMID: 22283778 Free PMC article. Review.
-
Common mechanisms regulating dark noise and quantum bump amplification in Drosophila photoreceptors.J Neurophysiol. 2013 Apr;109(8):2044-55. doi: 10.1152/jn.00001.2013. Epub 2013 Jan 30. J Neurophysiol. 2013. PMID: 23365183 Free PMC article.
-
Light-dependent translocation of visual arrestin regulated by the NINAC myosin III.Neuron. 2004 Jul 8;43(1):95-103. doi: 10.1016/j.neuron.2004.06.014. Neuron. 2004. PMID: 15233920
-
Regulation of arrestin translocation by Ca2+ and myosin III in Drosophila photoreceptors.J Neurosci. 2012 Jul 4;32(27):9205-16. doi: 10.1523/JNEUROSCI.0924-12.2012. J Neurosci. 2012. PMID: 22764229 Free PMC article.
-
Light-regulated translocation of signaling proteins in Drosophila photoreceptors.J Physiol Paris. 2006 Mar-May;99(2-3):133-9. doi: 10.1016/j.jphysparis.2005.12.010. Epub 2006 Feb 3. J Physiol Paris. 2006. PMID: 16458490 Free PMC article. Review.
Cited by
-
PDA (prolonged depolarizing afterpotential)-defective mutants: the story of nina's and ina's--pinta and santa maria, too.J Neurogenet. 2012 Jun;26(2):216-37. doi: 10.3109/01677063.2011.642430. Epub 2012 Jan 27. J Neurogenet. 2012. PMID: 22283778 Free PMC article. Review.
-
Invertebrate and vertebrate class III myosins interact with MORN repeat-containing adaptor proteins.PLoS One. 2015 Mar 30;10(3):e0122502. doi: 10.1371/journal.pone.0122502. eCollection 2015. PLoS One. 2015. PMID: 25822849 Free PMC article.
-
A role for the cytoplasmic DEAD box helicase Dbp21E2 in rhodopsin maturation and photoreceptor viability.J Neurogenet. 2012 Jun;26(2):177-88. doi: 10.3109/01677063.2012.692412. J Neurogenet. 2012. PMID: 22794106 Free PMC article.
-
Daily Regulation of Phototransduction, Circadian Clock, DNA Repair, and Immune Gene Expression by Heme Oxygenase in the Retina of Drosophila.Genes (Basel). 2018 Dec 21;10(1):6. doi: 10.3390/genes10010006. Genes (Basel). 2018. PMID: 30583479 Free PMC article.
-
Vitamin A Deficiency Alters the Phototransduction Machinery and Distinct Non-Vision-Specific Pathways in the Drosophila Eye Proteome.Biomolecules. 2022 Aug 6;12(8):1083. doi: 10.3390/biom12081083. Biomolecules. 2022. PMID: 36008977 Free PMC article.
References
-
- Ahmad ST, Natochin M, Artemyev NO, O'Tousa JE. The Drosophila rhodopsin cytoplasmic tail domain is required for maintenance of rhabdomere structure. FASEB J. 2007;21:449–455. - PubMed
-
- Arbeitman MN, Furlong EE, Imam F, Johnson E, Null BH, Baker BS, Krasnow MA, Scott MP, Davis RW, White KP. Gene expression during the life cycle of Drosophila melanogaster. Science. 2002;297:2270–2275. - PubMed
-
- Cagan RL, Ready DF. The emergence of order in the Drosophila pupal retina. Dev Biol. 1989;136:346–362. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/D007585/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- P20 RR017703/RR/NCRR NIH HHS/United States
- RR17703/RR/NCRR NIH HHS/United States
- BB/G006865/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- F32 EY006808/EY/NEI NIH HHS/United States
- EY13877/EY/NEI NIH HHS/United States
- EY06595/EY/NEI NIH HHS/United States
- R01 EY006808/EY/NEI NIH HHS/United States
- P30 EY012190/EY/NEI NIH HHS/United States
- R01 EY013877/EY/NEI NIH HHS/United States
- EY12190/EY/NEI NIH HHS/United States
- F32 EY006595/EY/NEI NIH HHS/United States
- EY06808/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases