

Song recognition learning and stimulus-specific weakening of neural responses in the avian auditory forebrain
- PMID: 20107117
- PMCID: PMC2853264
- DOI: 10.1152/jn.00885.2009
Song recognition learning and stimulus-specific weakening of neural responses in the avian auditory forebrain
Abstract
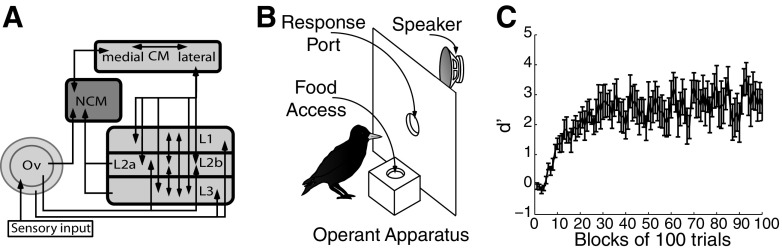
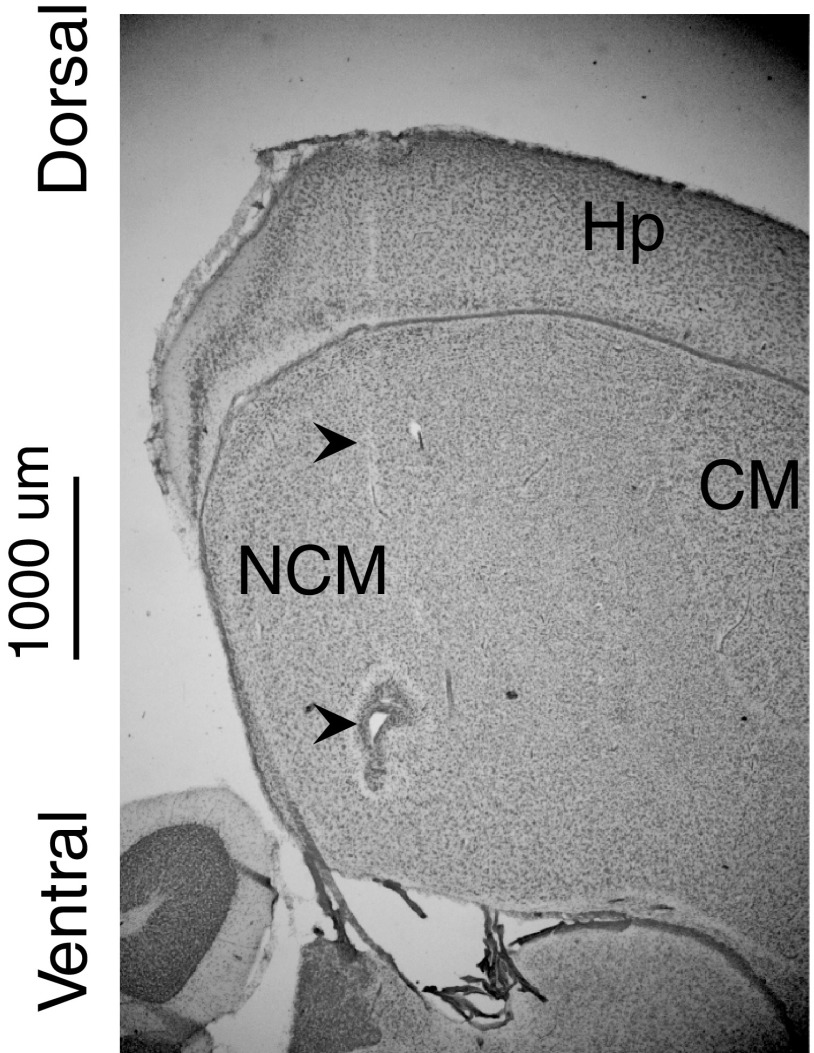
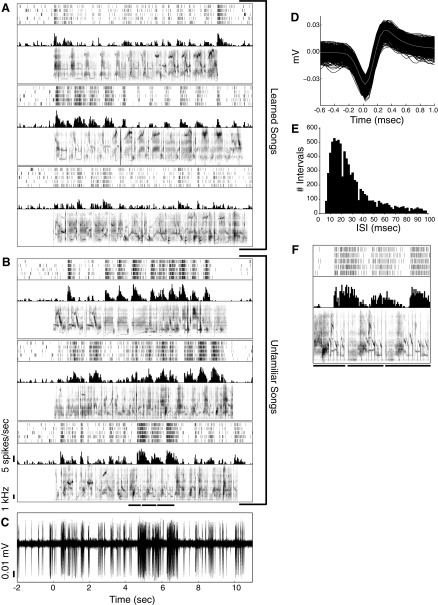
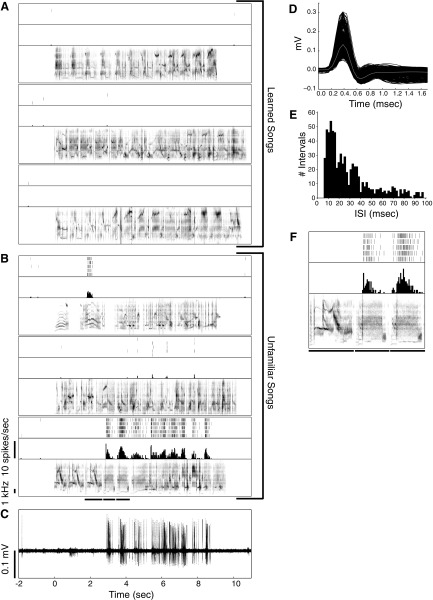
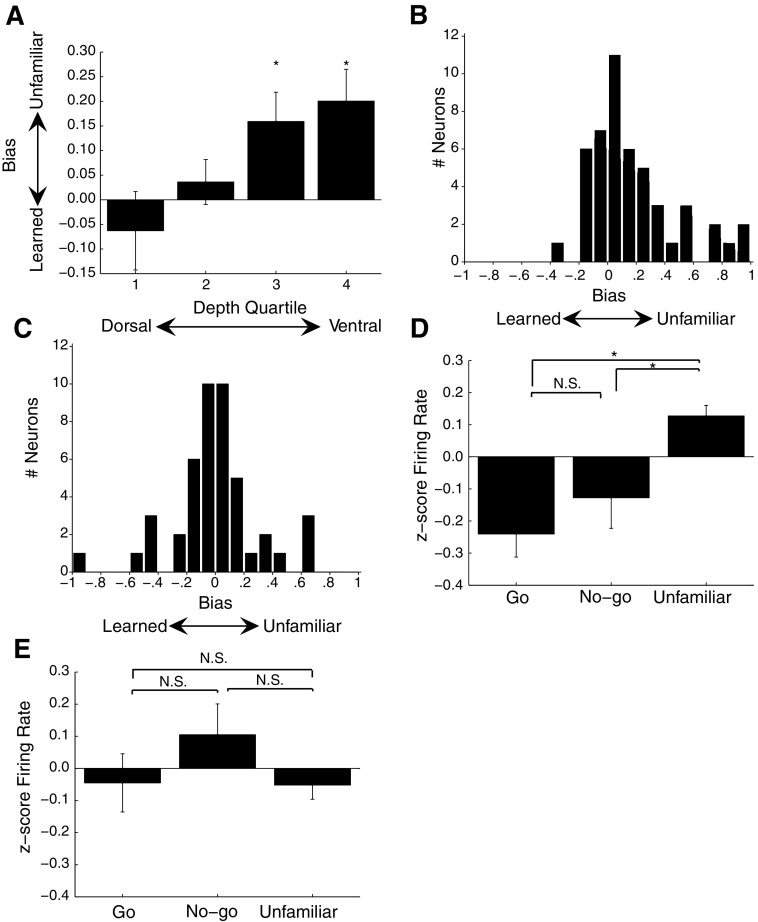
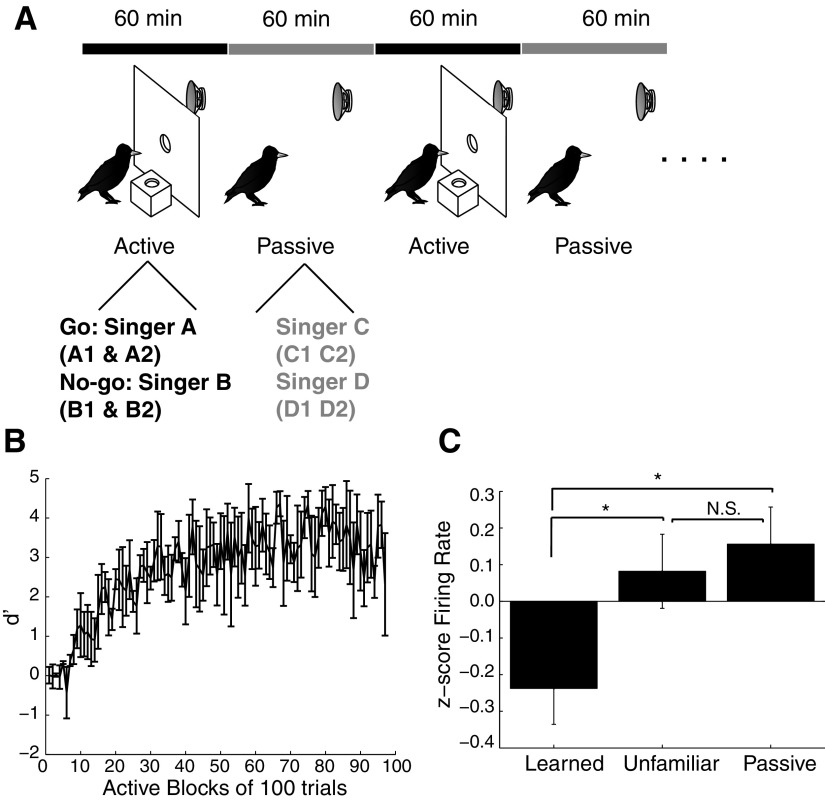
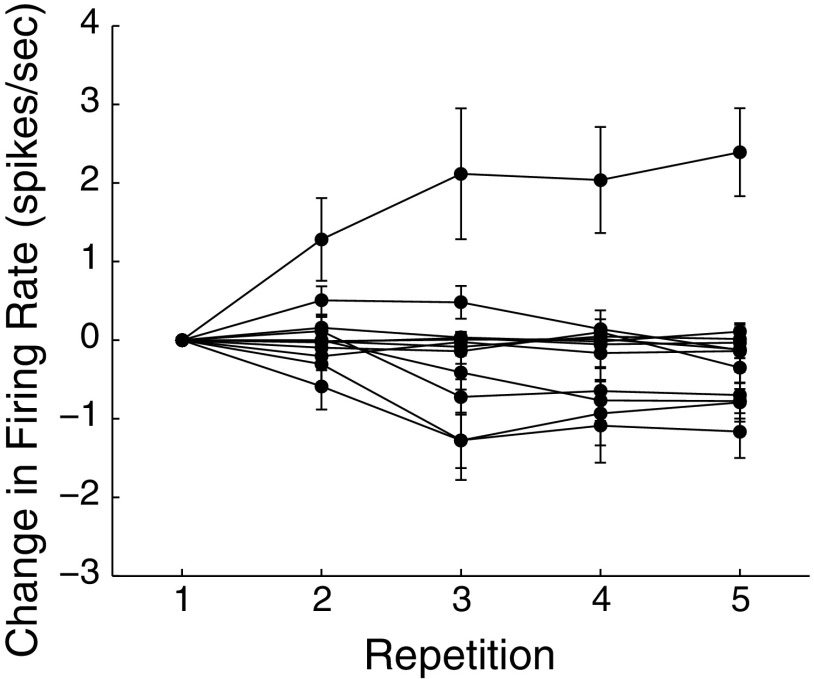
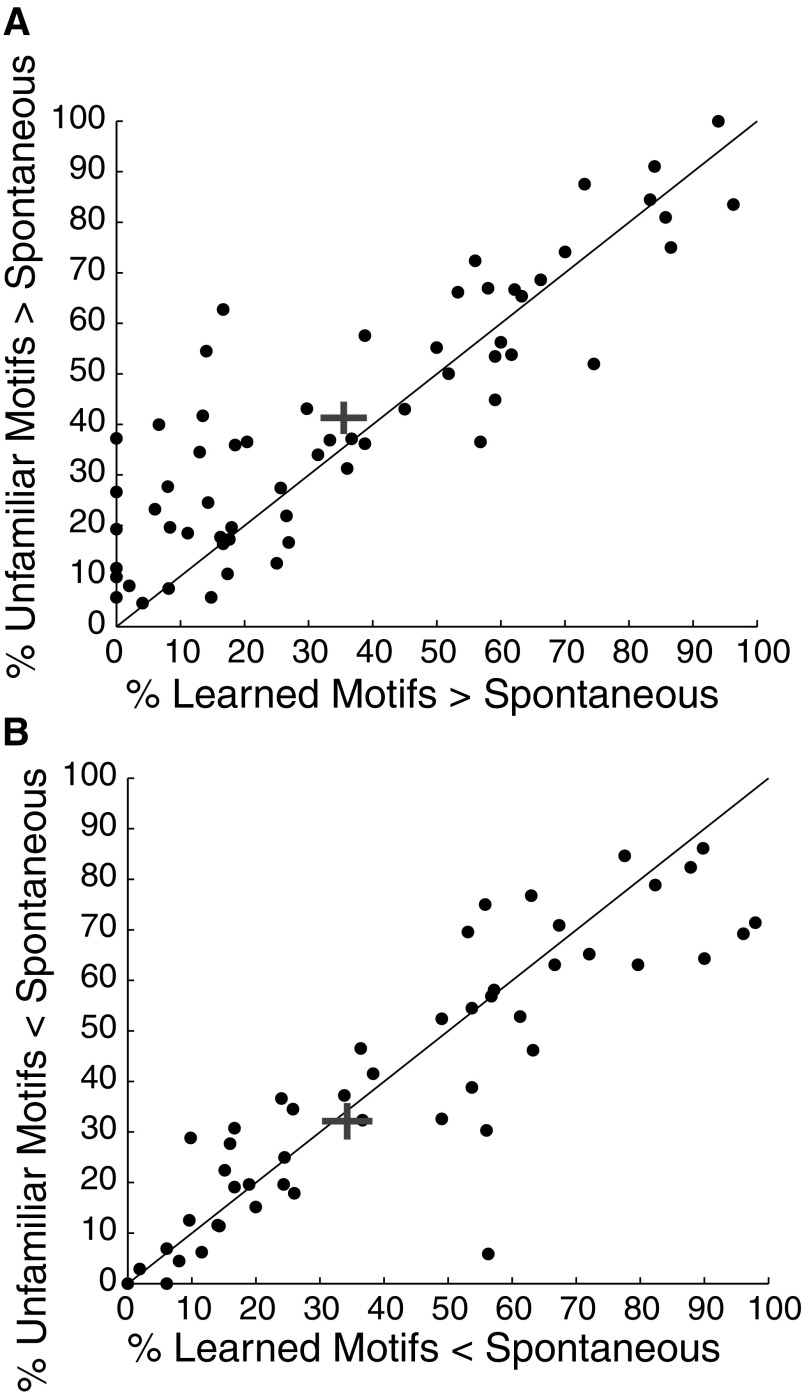
Learning typically increases the strength of responses and the number of neurons that respond to training stimuli. Few studies have explored representational plasticity using natural stimuli, however, leaving unknown the changes that accompany learning under more realistic conditions. Here, we examine experience-dependent plasticity in European starlings, a songbird with rich acoustic communication signals tied to robust, natural recognition behaviors. We trained starlings to recognize conspecific songs and recorded the extracellular spiking activity of single neurons in the caudomedial nidopallium (NCM), a secondary auditory forebrain region analogous to mammalian auditory cortex. Training induced a stimulus-specific weakening of the neural responses (lower spike rates) to the learned songs, whereas the population continued to respond robustly to unfamiliar songs. Additional experiments rule out stimulus-specific adaptation and general biases for novel stimuli as explanations of these effects. Instead, the results indicate that associative learning leads to single neuron responses in which both irrelevant and unfamiliar stimuli elicit more robust responses than behaviorally relevant natural stimuli. Detailed analyses of these effects at a finer temporal scale point to changes in the number of motifs eliciting excitatory responses above a neuron's spontaneous discharge rate. These results show a novel form of experience-dependent plasticity in the auditory forebrain that is tied to associative learning and in which the overall strength of responses is inversely related to learned behavioral significance.
Figures
Similar articles
-
Local inhibition modulates learning-dependent song encoding in the songbird auditory cortex.J Neurophysiol. 2013 Feb;109(3):721-33. doi: 10.1152/jn.00262.2012. Epub 2012 Nov 14. J Neurophysiol. 2013. PMID: 23155175 Free PMC article.
-
Emergence of learned categorical representations within an auditory forebrain circuit.J Neurosci. 2011 Feb 16;31(7):2595-606. doi: 10.1523/JNEUROSCI.3930-10.2011. J Neurosci. 2011. PMID: 21325527 Free PMC article.
-
Functional differences in forebrain auditory regions during learned vocal recognition in songbirds.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004 Dec;190(12):1001-10. doi: 10.1007/s00359-004-0556-x. Epub 2004 Sep 21. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004. PMID: 15449092
-
Neural correlates of learned song in the avian forebrain: simultaneous representation of self and others.Curr Opin Neurobiol. 2004 Aug;14(4):496-502. doi: 10.1016/j.conb.2004.06.004. Curr Opin Neurobiol. 2004. PMID: 15321071 Review.
-
Neural systems for individual song recognition in adult birds.Ann N Y Acad Sci. 2004 Jun;1016:282-302. doi: 10.1196/annals.1298.008. Ann N Y Acad Sci. 2004. PMID: 15313781 Review.
Cited by
-
Timing of perineuronal net development in the zebra finch song control system correlates with developmental song learning.Proc Biol Sci. 2018 Jul 18;285(1883):20180849. doi: 10.1098/rspb.2018.0849. Proc Biol Sci. 2018. PMID: 30051835 Free PMC article.
-
Active recognition enhances the representation of behaviorally relevant information in single auditory forebrain neurons.J Neurophysiol. 2013 Apr;109(7):1690-703. doi: 10.1152/jn.00461.2012. Epub 2013 Jan 9. J Neurophysiol. 2013. PMID: 23303858 Free PMC article.
-
Pattern-Induced Covert Category Learning in Songbirds.Curr Biol. 2015 Jul 20;25(14):1873-7. doi: 10.1016/j.cub.2015.05.046. Epub 2015 Jun 25. Curr Biol. 2015. PMID: 26119748 Free PMC article.
-
Increased bursting glutamatergic neurotransmission in an auditory forebrain area of the zebra finch (Taenopygia guttata) induced by auditory stimulation.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012 Sep;198(9):705-16. doi: 10.1007/s00359-012-0741-2. Epub 2012 Jul 3. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012. PMID: 22752655 Free PMC article.
-
Spike Train Coactivity Encodes Learned Natural Stimulus Invariances in Songbird Auditory Cortex.J Neurosci. 2021 Jan 6;41(1):73-88. doi: 10.1523/JNEUROSCI.0248-20.2020. Epub 2020 Nov 11. J Neurosci. 2021. PMID: 33177068 Free PMC article.
References
-
- Bakin JS, Weinberger NM. Classical conditioning induces CS-specific receptive field plasticity in the auditory cortex of the guinea pig. Brain Res 536: 271–286, 1990. - PubMed
-
- Bao S, Chang EF, Woods J, Merzenich MM. Temporal plasticity in the primary auditory cortex induced by operant perceptual learning. Nat Neurosci 7: 974–981, 2004. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources