

Review
doi: 10.1016/j.semcdb.2010.01.015.
Epub 2010 Jan 28.
Microtubule motors in eukaryotic spindle assembly and maintenance
Affiliations
- PMID: 20109569
- PMCID: PMC2927864
- DOI: 10.1016/j.semcdb.2010.01.015
Item in Clipboard
Review
Microtubule motors in eukaryotic spindle assembly and maintenance
Semin Cell Dev Biol.
2010 May.
Abstract
The spindle is a microtubule-based structure that facilitates chromosome segregation during mitosis and meiosis. Spindle assembly from dynamic microtubule building blocks is a major challenge for the dividing cell and a process that critically requires microtubule motors. In this review we focus on the mechanisms by which microtubule motors shape the spindle. Specifically, we address how motors are thought to move and arrange microtubules to form the characteristic bipolar morphology shared by all eukaryotic spindles as well as motor-dependent mechanisms of microtubule length regulation.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
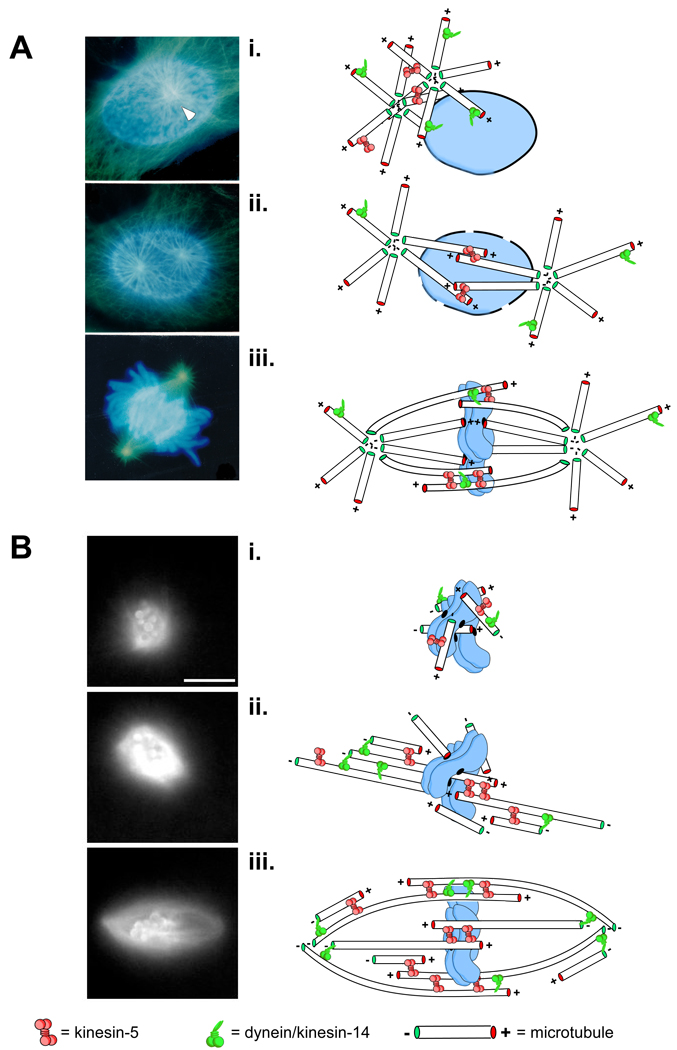
Pathways to spindle assembly are typically defined by the source of the microtubules used to construct the spindle. Some mitotic systems rely more heavily on one pathway or the other but spindle assembly likely involves some combination of both centrosome- and chromosome-nucleated microtubules. A, spindle assembly in cells from newt lungs is dominated centrosome-derived microtubules. Images are taken from fixed samples at different times during spindle assembly. In early prophase, astral microtubules (green) emanate from a pair of centrosomes (white arrowheads) clustered at a single locus on one side of the intact nucleus which bears condensed chromosomes, labeled in blue (i). The centrosomes are then forced apart by motor-dependent microtubule-microtubule sliding (e.g. kinesin-5 at sites of antiparallel overlap) as well as motors anchored on the nuclear envelope and at the cell cortex (ii). After nuclear envelop breakdown, centrosomally derived microtubules can search for and capture targets such as kinetochores and other microtubules ultimately adopting a spindle-like shape (iii). Acentrosomal spindle assembly, shown in B, is characterized by a burst of microtubule nucleation around chromatin (or chromatin-coated beads) that requires localized signals from the Ran-GTP and CPC pathways (see text; i). Monochromatic images were taken from a time-lapse recording of spindle assembly around chromatin-beads in Xenopus egg extracts doped with fluorescently labeled tubulin. Newly nucleated microtubules are arranged by sliding filament mechanisms eventually forming two prominent loci of focused minus ends, precursors to the spindle poles (ii). Eventually, microtubule polymer density equilibrates and the motors continue to shape the microtubule arrays until it achieves the bipolar, fusiform shape of a typical spindle (iii).
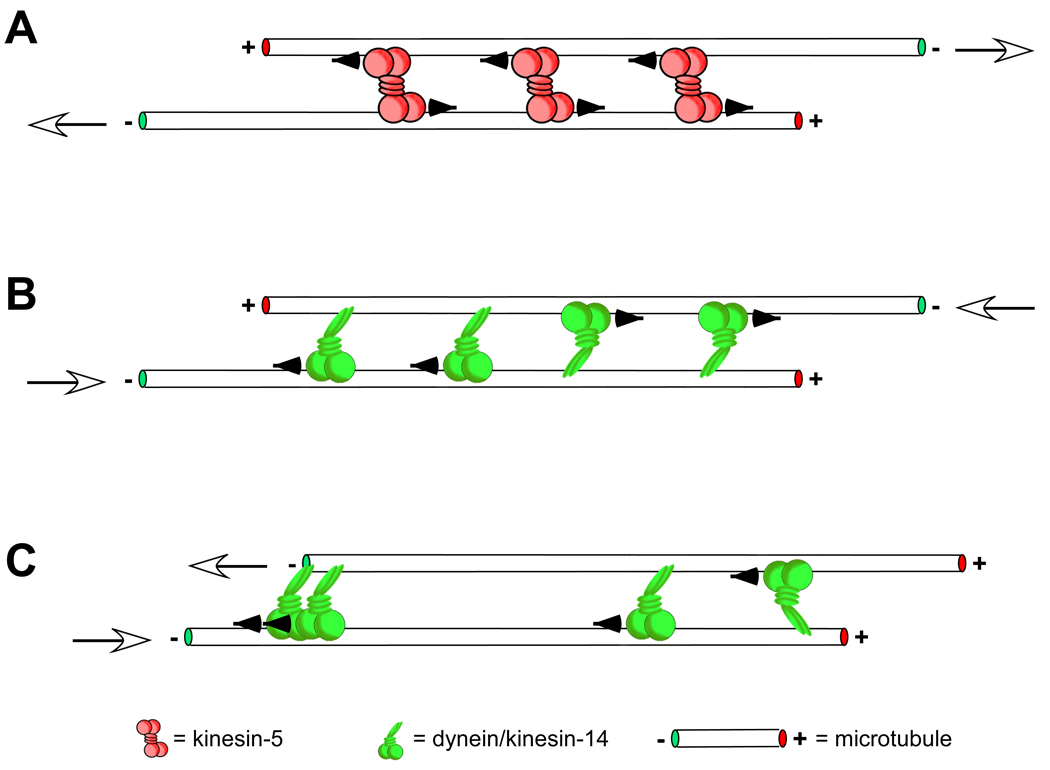
The orientation of cross-linked microtubules and the directional preference of the cross-linking motor dictate how sliding filament mechanisms contribute to spindle assembly and maintenance. In A, microtubules overlapped at their plus ends with an antiparallel orientation, which commonly occurs near the midzone of the spindle (see [20, 84, 85]), are pushed outward, toward the poles, by plus end-directed kinesin-5 motors. White arrows show the resulting direction of microtubule sliding. Conversely, minus end-directed motors such as dynein or kinesin-14 family members would act to pull the minus ends of the same microtubules together, sliding the microtubules inward, toward the spindle midzone ([20, 37]; B). A balance between these opposing forces is thought to contribute to achieving a steady-state spindle length. The other extreme of microtubule orientations is parallel alignment (C). In this geometry the effect of the motor on microtubule sliding depends on whether the motor “hangs on” once it reaches a microtubule end. A motor with bipolar symmetry like kinesin-5 would bind to and move processively along both the microtubules it cross-links without producing any relative sliding. In this way the motor may act to better align parallel microtubules along their lengths by “zippering” them up without sliding them [56]). For an asymmetric motor like dynein, which binds to microtubules via a static non-motor and steps along the other, the effect of stochastic binding of its non-motor end to either microtubule results in a net force of zero (e.g. the two motors on the right-hand side of the cross-linked microtubules have equal and opposite effects on microtubule sliding, producing no net sliding or force). However, a higher motor binding affinity to microtubule minus ends would allow for sliding and end alignment (for an excellent treatment of these mechanisms please see [17]).
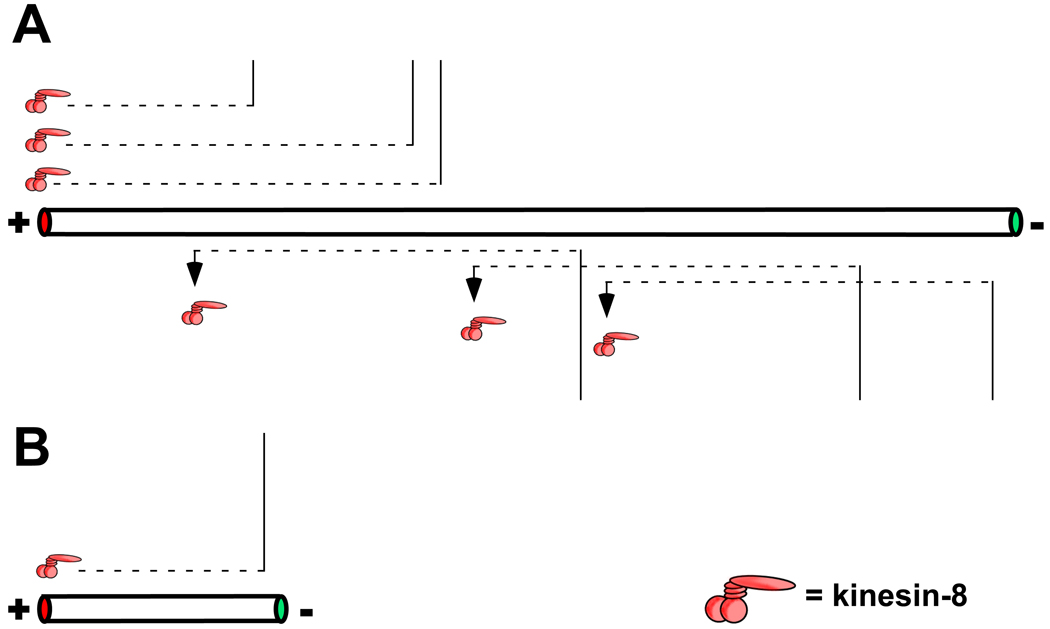
The ability of a motor-depolymerases to affect microtubule length in a length-dependent manner is derived from the distance the motor can travel during its association with the microtubule track. For a processive motor, this distance is proportional the velocity of the motor and the amount of time it spends attached to the microtubule [71]. For a motor that relies on lattice based diffusion, such as MCAK, this is likely dependent on diffusion length which is proportional to the amount of time the motor dwells on the microtubule and its diffusion coefficient [69]. The diffusion length for MCAK is less than 1 µm while that for the processive kinesin-8 is ~ 8 µm. Microtubules longer than the motor’s typical travel length will collect the most motors at their tips, increasing the chance of a catastrophe (A). Motors that bind the microtubule too far from the ends will fall off before reaching them and not influence catastrophe frequency. In B, shorter microtubules will not collect as many motors and therefore will have fewer motors at their tips reducing the chance of catastrophe.
References
-
- Walczak CE, Heald R. Mechanisms of mitotic spindle assembly and function. Int Rev Cytol. 2008;265:111–158. - PubMed
-
- Mitchison T, Kirschner M. Dynamic instability of microtubule growth. Nature. 1984;312:237–242. - PubMed
-
- Kirschner M, Mitchison T. Beyond self-assembly: from microtubules to morphogenesis. Cell. 1986;45:329–342. - PubMed
-
- Wollman R, Cytrynbaum EN, Jones JT, Meyer T, Scholey JM, Mogilner A. Efficient chromosome capture requires a bias in the 'search-and-capture' process during mitotic-spindle assembly. Curr Biol. 2005;15:828–832. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
