

Review
doi: 10.1038/nature08912.
Towards building a chromosome segregation machine
Affiliations
- PMID: 20110988
- PMCID: PMC2879044
- DOI: 10.1038/nature08912
Item in Clipboard
Review
Towards building a chromosome segregation machine
Nature.
.
Abstract
All organisms, from bacteria to humans, face the daunting task of replicating, packaging and segregating up to two metres (about 6 x 10(9) base pairs) of DNA when each cell divides. This task is carried out up to a trillion times during the development of a human from a single fertilized cell. The strategy by which DNA is replicated is now well understood. But when it comes to packaging and segregating a genome, the mechanisms are only beginning to be understood and are often as variable as the organisms in which they are studied.
Figures

a, Polymer repulsion in a confined space. Chromosomal DNA is confined within the boundaries of a cell. DNA is depicted here as tension blobs, which represent the random coil of DNA within the length scale at which thermal forces randomize the conformation of the DNA polymer. Entropy drives fluctuation of each chain (dark and light) to adopt a random coil. Segregation occurs as a result of the entropic repulsion between two chains in a confined space. This mechanism has been proposed to operate in prokayotes and might contribute to segregation in eukaryotes that undergo closed mitosis (such as fungi and some protists). b, Polymer growth. The growth of an actin-like polymer (blue) generates force, which propels plasmid DNA (depicted as curved DNA helices) to the opposite sides of a prokaryotic cell. c, Polymer brushes. Multiple polymers can be concentrated on opposite sides of the cell wall, analogous to a brush in which short, flexible bristles are attached to a stiff handle. Polymer physicists use this type of strategy to switch force rapidly from attraction to repulsion. In the bacterium Caulobacter crescentus, polymers of PopZ form a network attached to opposite sides of the bacterial cell wall and anchor the two circular DNA chromosomes to effect chromosome segregation. d, Mitotic spindle. Antiparallel arrays of dynamic microtubules (green rods) extend from microtubule-organizing centres (green ovals) at opposite poles of the eukaryotic cell. These microtubules attach to chromosomes (red) (which are wrapped around histone proteins to form nucleosomes, green) by way of the kinetochore (blue). Ring-like complexes (pink) hold replicated sister chromatids together. The spindle microtubules provide an outward force (towards the spindle pole) that acts against the inward force emanating from the linkage between sister chromatids and drives segregation.

Regions of a chromosome can be unravelled, forming a loop, either to allow specialized functions (such as cell division) or in specific cell types or at specific developmental stages. Examples of DNA extrusion from the axis of the chromosome are shown. a, DNA looping was first observed in squash preparations of salamander eggs under the light microscope, by the embryologist Oskar Hertwig in the early 1900s. A single egg is shown here. Scale bar, 50 μm. b, Paulson and Laemmli found DNA loops when examining chromosome spreads in isolated mammalian cells. In the electron micrograph of this cell in metaphase, loops emanate from the protein-rich chromosome scaffold (the darker stained, X-shaped structure towards the bottom of the image); the inset shows a whole metaphase chromosome, highlighting its similarity to the isolated chromosome scaffold. Scale bars, 2 μm. (Image reproduced, with permission, from ref. .) c, DNA loops were also found on a single meiotic bivalent chromosome from the oocyte of the newt Triturus viridescens, by using light microscopy. These are sites of intense transcriptional activity. Each lateral loop represents a single DNA duplex. Scale bar, 15 μm. (Image reproduced, with permission, from ref. .)d, And loops were observed in DNA undergoing transcription when heat-shock loci in the fly Chironomus tentans were examined by using light microscopy. The depiction of the findings shows chromosomal DNA (red) as a strand that forms loops (centre) but elsewhere is wrapped tightly in bundles (green; proposed by the authors to be nucleosomes). e, DNA looping was first proposed to occur at the kinetochore in mammals. Plates of microtubule-binding segments are tandemly repeated and interspersed with linker segments. On microtubule attachment, microtubule-binding segments loop out from the chromosome axis, forming a cluster of repeats on the surface of the chromosome, as depicted for one loop of centromeric DNA in f. f, DNA looping was later proposed to occur in Saccharomyces cerevisiae also. The single pericentromeric loop is comparable to multiple loops of linker segments and microtubule-binding segments (e) that form on microtubule attachment in mammalian cells. The kinetochore (not shown here) forms at the junction between microtubules (green rods) and the unique nucleosome (blue) at the apex of the pericentromeric loop.

a, SMC proteins assemble into complexes that adopt a ring-like conformation. The backbone of the ring is formed by the SMC proteins themselves (MukB in bacteria; Smc2 and Smc4 in Saccharomyces cerevisiae condensin, and Smc1 and Smc3 in S. cerevisiae cohesin). In eukaryotes, the SMC monomer is folded in an antiparallel coiled coil. At one end (top of molecule as depicted), the two monomers associate to form a hinge, and at the other end is an ATP-binding head domain. Closure of the ring at the head domain is carried out by proteins known as kleisins, including Scc1 (also known as Mcd1) and Brn1. Each dimer is associated with additional proteins (for example Ysc4, Ycg1, Scc3 (also known as Irr1), Rad61 and Pds5) at the head domain to form a functional complex in vivo. In bacteria, the SMC coiled coils are bound by ScpA and ScpB. b, Cohesin rings (pink) and condensin rings (blue) are depicted linking chromatids into a network (that is creating links within a chromatid, such as those that allow DNA looping) and, for cohesin, holding sister chromatids. c, A model for how cohesin and condensin rings may contribute directly to the elastic properties of the centromere and/or chromosome. Protein rings that function as slip rings (or molecular pulleys) provide a mechanism to distribute tension from one location to the entire network, and cohesin (pink) and condensin (blue) have the physical attributes to function as slip rings and regulate centromere elasticity.
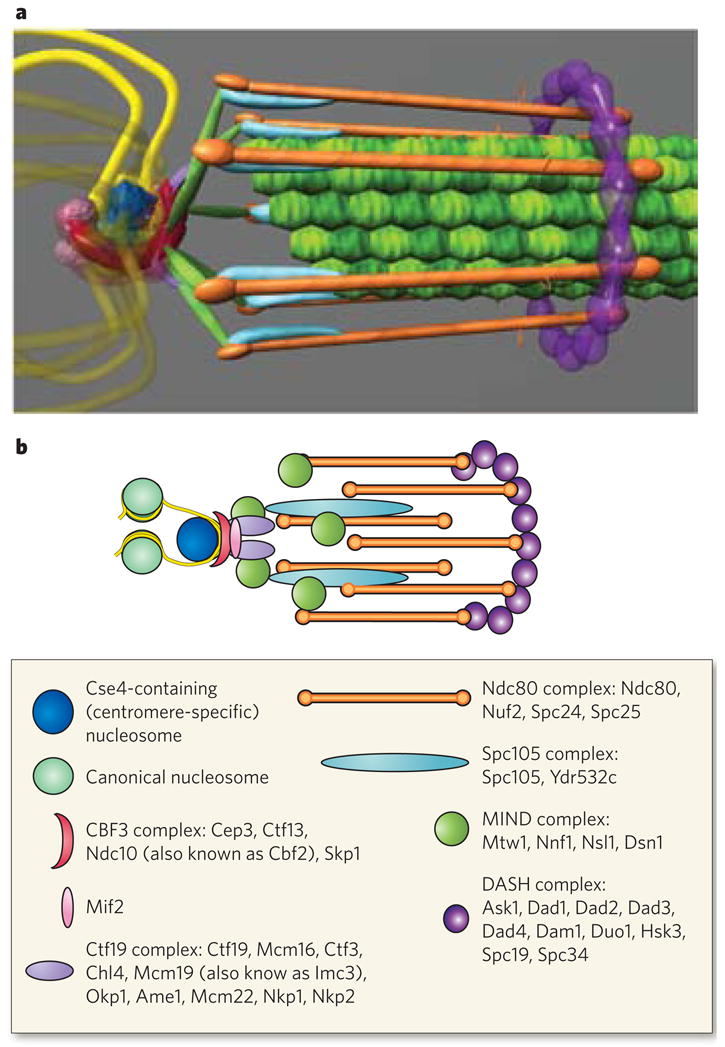
The structure of the kinetochore is depicted in two ways. a, This structure reflects the average positions of kinetochore proteins within the S. cerevisiae kinetochore measured by invivo super-resolution microscopy. The positions of microtubule-binding proteins with respect to the microtubule plus end are key to understanding the mechanism of force generation at the kinetochore. The interface between the microtubule-binding side of the kinetochore and the chromatin-binding side in S. cerevisiae, as well as in multicellular eukaryotes, remains poorly characterized. From the left, the DNA (yellow strands) is wrapped in a positive supercoil around a histone core that contains the centromere-specific H3 variant (dark blue). The DNA-binding region of the kinetochore is formed by the CBF3 complex (dark pink). An additional DNA-binding protein, Mif2, is shown proximal to centromeric DNA (light pink sphere). From the right, the microtubules (green filaments) are surrounded by the DASH complex (purple ring), whose constituents are abundant enough to form a ring. The main microtubule-binding sites within the kinetochore are in the Ndc80 complex (orange rods). There are several linker complexes: the Spc105 complex is depicted in light blue, and the MIND complex in green; the Ctf19 complex is shown as small purple linkers between the CBF3 and MIND complexes. b, This illustration is a schematic representation of the structure in a. Data are based on the hydrodynamic properties, protein number and spatial position of the members of each protein complex. The structure assumes a symmetrical arrangement of kinetochore protein complexes around the microtubule lattice (green in a). The known components of the various protein complexes are listed in the box.

Two types of mechanism have been proposed: biased diffusion (a) and forced walk (b, c). a, Microtubules can be persistently attached to lattice-binding proteins such as the Ndc80 complex (Fig. 4) through many diffusive couplers (for example, flexible, positively charged amino acids at the amino terminus of the Ndc80 subunit of the Ndc80 complex94,95) attached at different distances from the plus end. Thermal diffusion is depicted as random fluctuations of the microtubule lattice (double-headed arrows). Depolymerization at the microtubule plus end provides a bias to the diffusive movements while attachment can be maintained. b, The discovery of DASH rings and subsequent in vitro studies led to the idea that ring couplers (such as the DASH complex) are involved in force generation. Energy from microtubule depolymerization drives the rings towards the microtubule minus end (curved arrows). c, Subsequently, ultrastructural study of the vertebrate kinetochore uncovered the presence of fibrillar couplers (pink) that attach to peeling microtubule protofilaments at specific locations. This discovery led to the idea that fibrillar couplers can themselves be force generators. The mechanisms proposed in a, b and c are all remain valid. d, Finally, when the plus end of a microtubule polymerizes against a rigid barrier, such as the cell wall, this can also generate forces, albeit smaller forces, but these forces are not thought to have a significant role in kinetochore motility.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
