

Review
doi: 10.1038/nature08908.
Cell mechanics and the cytoskeleton
Affiliations
- PMID: 20110992
- PMCID: PMC2851742
- DOI: 10.1038/nature08908
Item in Clipboard
Review
Cell mechanics and the cytoskeleton
Nature.
.
Abstract
The ability of a eukaryotic cell to resist deformation, to transport intracellular cargo and to change shape during movement depends on the cytoskeleton, an interconnected network of filamentous polymers and regulatory proteins. Recent work has demonstrated that both internal and external physical forces can act through the cytoskeleton to affect local mechanical properties and cellular behaviour. Attention is now focused on how cytoskeletal networks generate, transmit and respond to mechanical signals over both short and long timescales. An important insight emerging from this work is that long-lived cytoskeletal structures may act as epigenetic determinants of cell shape, function and fate.
Figures
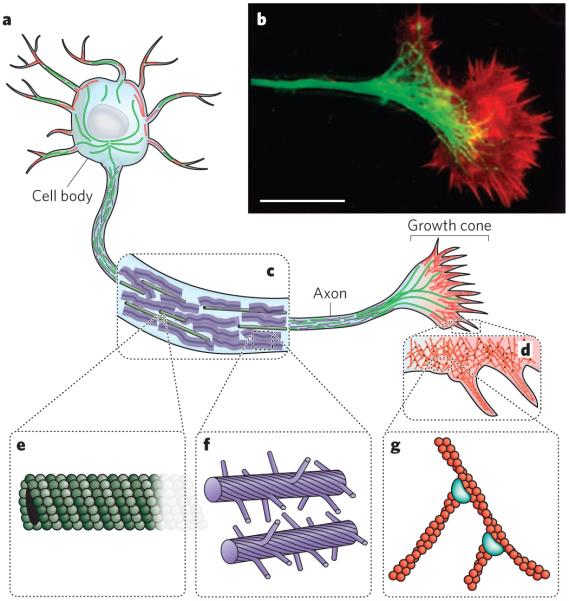
The cytoskeleton of eukaryotic cells provides structure and organization, resists and transmits stresses, and drives shape change and movement. a, Neurons are specialized eukaryotic cells that extend long processes to form connections in the nervous system. Like other eukaryotic cells, neurons have a cytoskeleton that consists of three main polymers: microtubules (green), intermediate filaments (purple) and actin filaments (red). b, A fluorescence micrograph of the neuronal growth cone, which migrates in response to chemical cues during the development of the nervous system, is shown. Microtubules (green) emanate from the axon, and actin-filament networks (red) form sheet-like structures and filopodial protrusions at the leading edge. Scale bar, 20 μm. (Image reproduced, with permission, from ref. .) c, The neuronal axon is a long membrane-bounded extension, in which neurofilaments (a class of intermediate filament in neurons) form a structural matrix that embeds microtubules, which transport materials from the cell body to the axon terminals at the synapse. d, The growth cone contains dendritic actinfilament networks and parallel actin-filament filopodia. e, Microtubules consist of 13 protofilaments of tubulin dimers arranged in a hollow tube. f, Neurofilaments have flexible polymer arms that repel neighbouring neurofilaments and determine the radius of the axon. g, Actin filaments are arranged into networks. These networks can have many architectures, including the branched structures depicted here, which are formed by the Arp2/3 complex (blue). The diameters of microtubules, intermediate filaments and actin filaments are within a factor of three of each other; the diagrams in e, f and g are drawn approximately to scale. But the relative flexibilities of these polymers differ markedly, as indicated by their persistence lengths: from least to most flexible, microtubules (5,000 μm), actin filaments (13.5 μm) and intermediate filaments (0.5 μm).
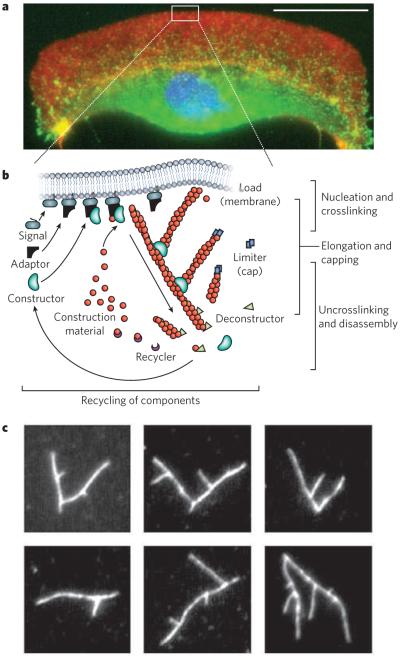
Long-range order of the cytoskeleton is generated by simple rules for network assembly and disassembly. a, A fluorescence micrograph of a fish keratocyte is shown (with the nucleus in blue). Motile cells such as these form branched actin-filament networks (red) at their leading edge, and these branched networks generate protrusions. Together with coordinated adhesions to a surface (indicated by vinculin, green) and myosin-driven retraction, the protrusions lead to directed movement. Scale bar, 15 μm. (Image courtesy of M. van Duijn, Univ. California, Berkeley.) b, There are three basic steps involved in the assembly of protrusive, branched actin-filament networks: filament elongation; nucleation and crosslinking of new filaments from filaments close to the membrane; and capping of filaments. Disassembly of the network involves a separate set of proteins that severs the filaments and recycles the subunits. c, The branching of actin filaments can be reconstituted in vitro with soluble proteins, generating various branched structures such as those in these fluorescence micrographs of labelled actin (white). (Images courtesy of O. Akin, Univ. California, San Francisco.)
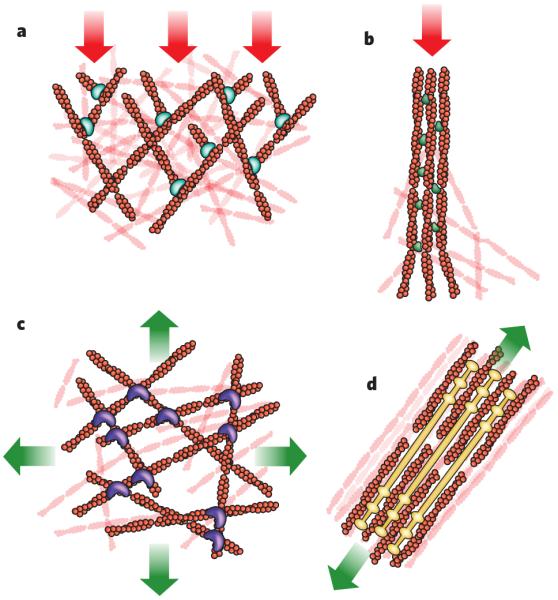
The cytoskeleton forms structures that have a wide variety of architectures and are associated with different types of cellular force. Shown are four structures generated by actin filaments and the stresses typically encountered by these structures (red arrows, compression; green arrows, tension). a, Branched actin-filament networks push against the plasma membrane and external barriers as they generate protrusions, thereby encountering an inward force of compression. b, Filaments bundled into filopodia also generate protrusive forces as they extend from the cell body, encountering similar compressive force. In this case, the linker molecule is the bundling protein fascin. c, Cortical networks (that is, non-aligned networks), such as this one involving filamin as a crosslinker, form below the plasma membrane and carry tension loads in multiple directions. d, Stress fibres form from bundled actin filaments, shown here associated with filaments of myosin, and generate tension against cell adhesions to the extracellular matrix. The illustrations are based on micrographs from refs - (a–d, respectively).
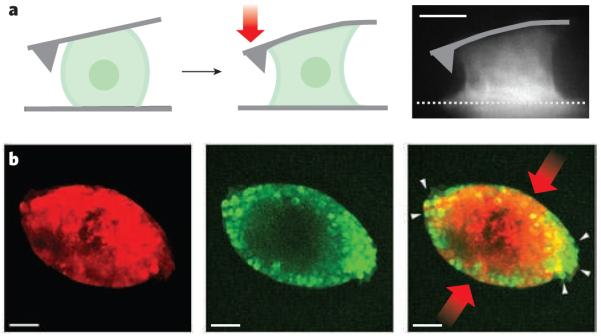
a, Single cells can exert large contractile forces that affect their shape. An osteosarcoma cell attached between the cantilever of an atomic force microscope and a surface can exert contractile forces (red arrow) of more than 100 nN (depicted diagramatically from left panel to centre panel and in a fluorescence micrograph, right panel). Actin-filament structures (white), including contractile stress fibres spanning the upper and lower surfaces, are generated in the contracting osteosarcoma cell (right). Scale bar, 10 μm. (Image reproduced, with permission, from ref. .) b, Mechanical stress on cancer cells in three-dimensional tumour spheroids changes their growth. There are fewer proliferating cells (green) in tumour spheroids (red) at the regions of highest compressive stress (red arrows) than at the regions of low stress (white arrows). The image on the right is an overlay of the centre and left images. Scale bar, 50 μm. (Images reproduced, with permission, from ref. .)
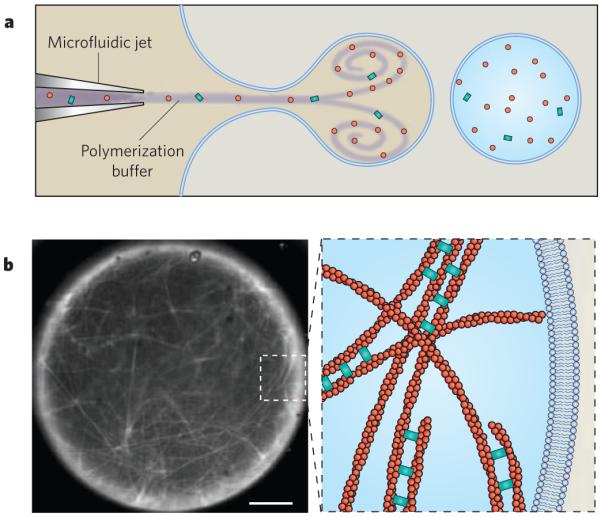
The reconstitution of cytoskeletal structures is a key method for understanding how functional behaviour emerges from discrete components. In this example, actin filaments were nucleated from beads coated with the nucleation-promoting factor ActA (not shown) and were then crosslinked by fascin inside a unilamellar lipid vesicle. a, Purified proteins were loaded into a vesicle by a microfluidic encapsulation technique that allows the dynamics of filament assembly to be observed immediately after encapsulation. b, The micrograph (left) shows fluorescently labelled actin filaments (white) that have polymerized inside the vesicle and have assembled into a fascin-crosslinked network. Scale bar, 5 μm. The diagram (right) is a schematic depiction of the actin-filament network present in the inset box of the micrograph. (Image courtesy of D. Richmond, S. Hansen and M. Zanic, Physiology Course, Marine Biological Laboratory, Woods Hole, Massachusetts.)
References
-
- Weiss PA. In: The Molecular Control of Cellular Activity. Allen JM, editor. McGraw-Hill; 1961. pp. 1–72.
-
- dos Remedios CG, et al. Actin binding proteins: regulation of cytoskeletal microfilaments. Physiol. Rev. 2003;83:433–473. - PubMed
-
- Bieling P, et al. Reconstitution of a microtubule plus-end tracking system in vitro. Nature. 2007;450:1100–1105. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
