

Crosstalk between C/EBPbeta phosphorylation, arginine methylation, and SWI/SNF/Mediator implies an indexing transcription factor code
- PMID: 20111005
- PMCID: PMC2845275
- DOI: 10.1038/emboj.2010.3
Crosstalk between C/EBPbeta phosphorylation, arginine methylation, and SWI/SNF/Mediator implies an indexing transcription factor code
Abstract
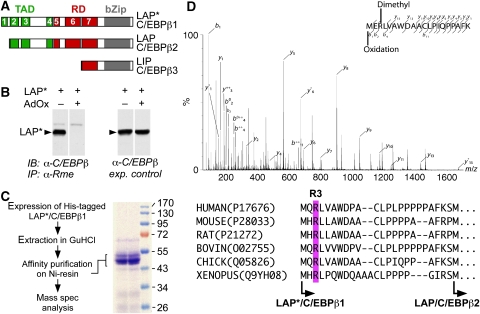
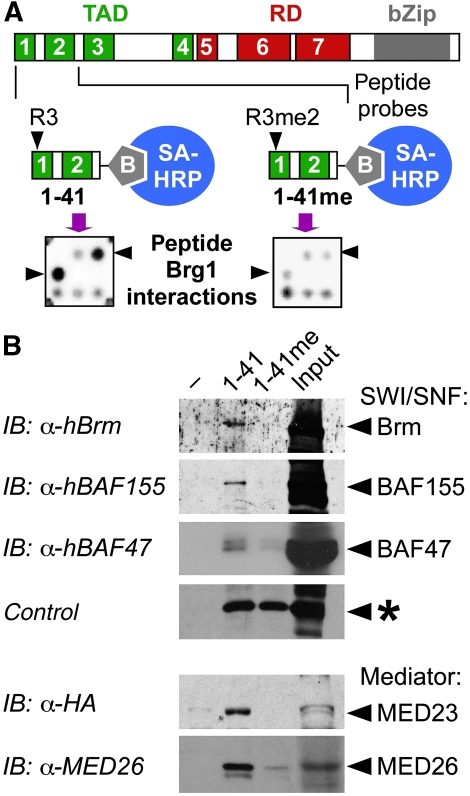
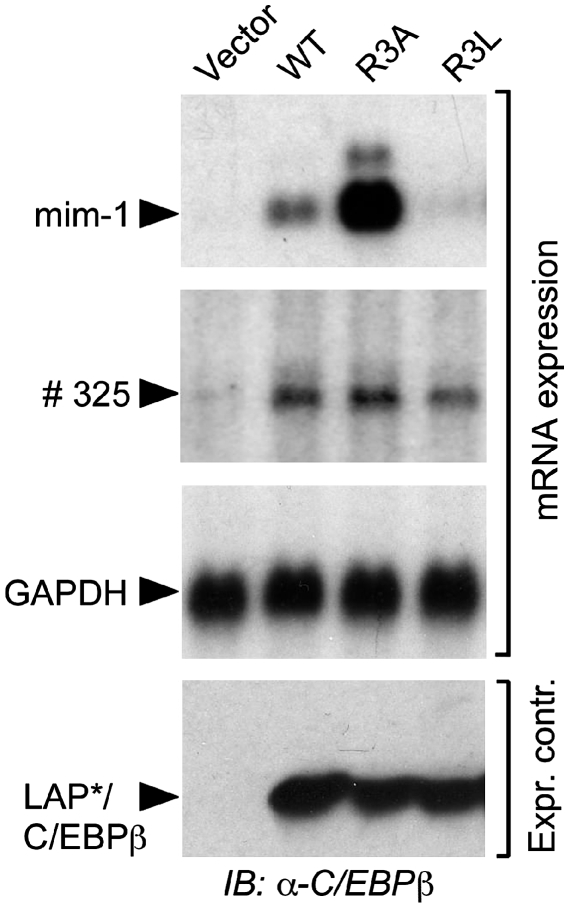
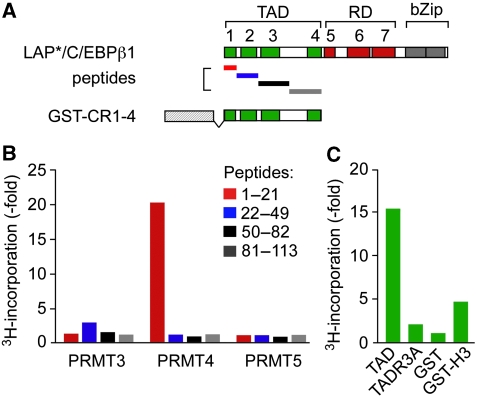
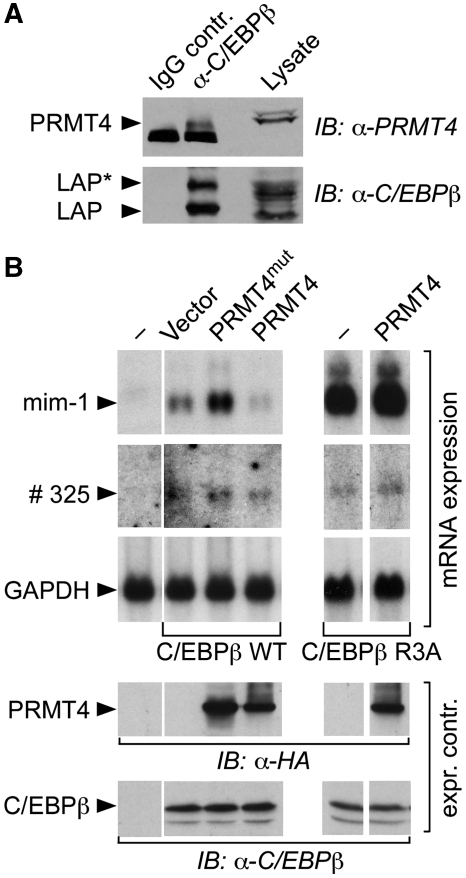
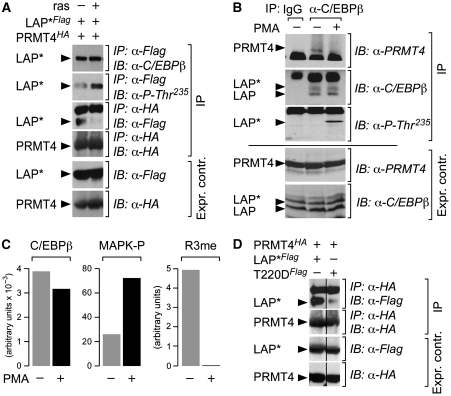
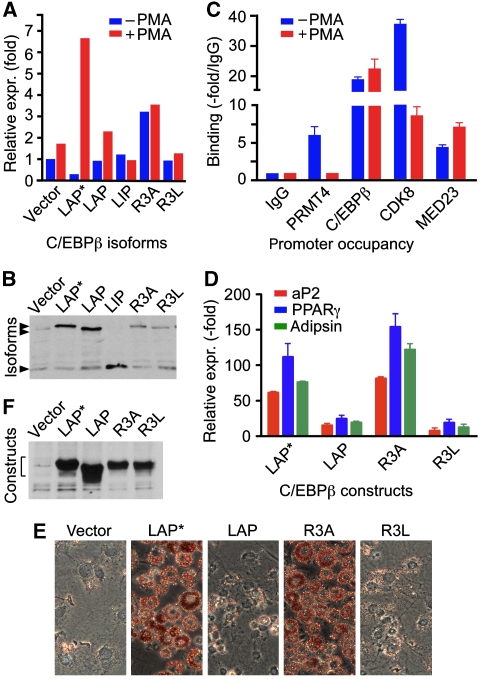
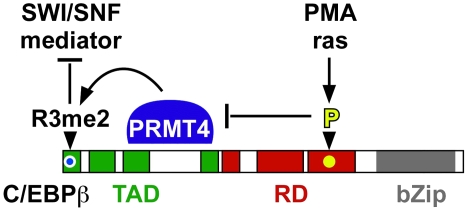
Cellular signalling cascades regulate the activity of transcription factors that convert extracellular information into gene regulation. C/EBPbeta is a ras/MAPkinase signal-sensitive transcription factor that regulates genes involved in metabolism, proliferation, differentiation, immunity, senescence, and tumourigenesis. The protein arginine methyltransferase 4 PRMT4/CARM1 interacts with C/EBPbeta and dimethylates a conserved arginine residue (R3) in the C/EBPbeta N-terminal transactivation domain, as identified by mass spectrometry of cell-derived C/EBPbeta. Phosphorylation of the C/EBPbeta regulatory domain by ras/MAPkinase signalling abrogates the interaction between C/EBPbeta and PRMT4/CARM1. Differential proteomic screening, protein interaction studies, and mutational analysis revealed that methylation of R3 constraines interaction with SWI/SNF and Mediator complexes. Mutation of the R3 methylation site alters endogenous myeloid gene expression and adipogenic differentiation. Thus, phosphorylation of the transcription factor C/EBPbeta couples ras signalling to arginine methylation and regulates the interaction of C/EBPbeta with epigenetic gene regulatory protein complexes during cell differentiation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
A functional N-terminal domain in C/EBPβ-LAP* is required for interacting with SWI/SNF and to repress Ric-8B gene transcription in osteoblasts.J Cell Physiol. 2014 Oct;229(10):1521-8. doi: 10.1002/jcp.24595. J Cell Physiol. 2014. PMID: 24585571 Free PMC article.
-
Novel mechanism of negative regulation of 1,25-dihydroxyvitamin D3-induced 25-hydroxyvitamin D3 24-hydroxylase (Cyp24a1) Transcription: epigenetic modification involving cross-talk between protein-arginine methyltransferase 5 and the SWI/SNF complex.J Biol Chem. 2014 Dec 5;289(49):33958-70. doi: 10.1074/jbc.M114.583302. Epub 2014 Oct 16. J Biol Chem. 2014. PMID: 25324546 Free PMC article.
-
The Ric-8B gene is highly expressed in proliferating preosteoblastic cells and downregulated during osteoblast differentiation in a SWI/SNF- and C/EBPbeta-mediated manner.Mol Cell Biol. 2011 Jul;31(14):2997-3008. doi: 10.1128/MCB.05096-11. Epub 2011 May 23. Mol Cell Biol. 2011. PMID: 21606199 Free PMC article.
-
CARM1/PRMT4: Making Its Mark beyond Its Function as a Transcriptional Coactivator.Trends Cell Biol. 2021 May;31(5):402-417. doi: 10.1016/j.tcb.2020.12.010. Epub 2021 Jan 20. Trends Cell Biol. 2021. PMID: 33485722 Review.
-
Interplay between chromatin remodelers and protein arginine methyltransferases.J Cell Physiol. 2007 Nov;213(2):306-15. doi: 10.1002/jcp.21180. J Cell Physiol. 2007. PMID: 17708529 Review.
Cited by
-
A differential proteome screening system for post-translational modification-dependent transcription factor interactions.Nat Protoc. 2011 Mar;6(3):359-64. doi: 10.1038/nprot.2011.303. Epub 2011 Feb 24. Nat Protoc. 2011. PMID: 21372816
-
PRISMA and BioID disclose a motifs-based interactome of the intrinsically disordered transcription factor C/EBPα.iScience. 2021 Jun 4;24(6):102686. doi: 10.1016/j.isci.2021.102686. eCollection 2021 Jun 25. iScience. 2021. PMID: 34189442 Free PMC article.
-
Carm1 regulates Pax7 transcriptional activity through MLL1/2 recruitment during asymmetric satellite stem cell divisions.Cell Stem Cell. 2012 Sep 7;11(3):333-45. doi: 10.1016/j.stem.2012.07.001. Epub 2012 Aug 2. Cell Stem Cell. 2012. PMID: 22863532 Free PMC article.
-
Pro-inflammatory cytokine induction of 11β-hydroxysteroid dehydrogenase type 1 in A549 cells requires phosphorylation of C/EBPβ at Thr235.PLoS One. 2013 Sep 26;8(9):e75874. doi: 10.1371/journal.pone.0075874. eCollection 2013. PLoS One. 2013. PMID: 24086653 Free PMC article.
-
C/EBP maintains chromatin accessibility in liver and facilitates glucocorticoid receptor recruitment to steroid response elements.EMBO J. 2013 May 29;32(11):1568-83. doi: 10.1038/emboj.2013.106. Epub 2013 May 10. EMBO J. 2013. PMID: 23665916 Free PMC article.
References
-
- Baer M, Johnson PF (2000) Generation of truncated C/EBPbeta isoforms by in vitro proteolysis. J Biol Chem 275: 26582–26590 - PubMed
-
- Bartel RL, Borchardt RT (1984) Effects of adenosine dialdehyde on S-adenosylhomocysteine hydrolase and S-adenosylmethionine-dependent transmethylations in mouse L929 cells. Mol Pharmacol 25: 418–424 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
