

Kinesin-1/Hsc70-dependent mechanism of slow axonal transport and its relation to fast axonal transport
- PMID: 20111006
- PMCID: PMC2829163
- DOI: 10.1038/emboj.2009.389
Kinesin-1/Hsc70-dependent mechanism of slow axonal transport and its relation to fast axonal transport
Abstract
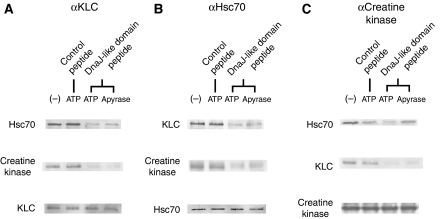
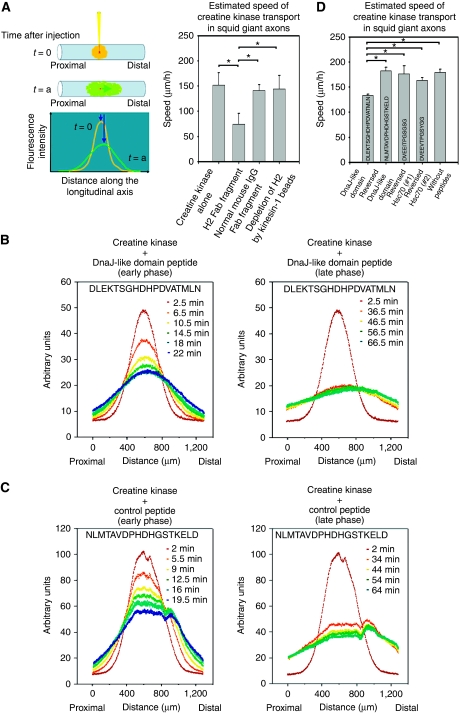
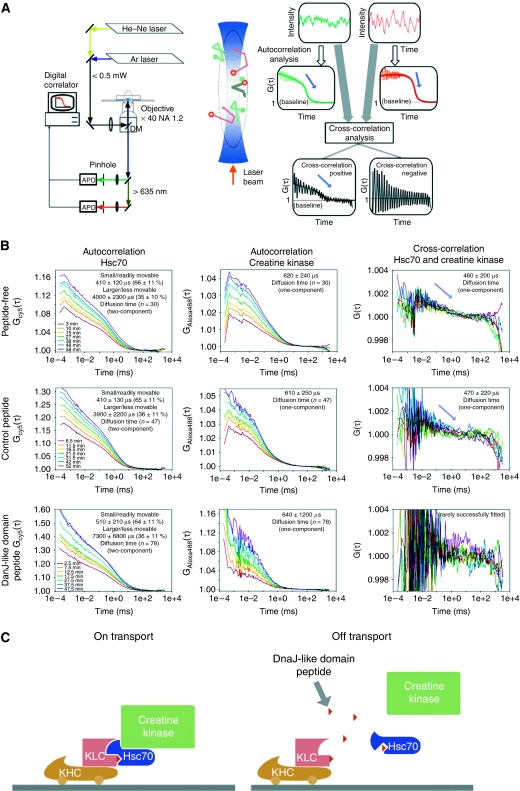
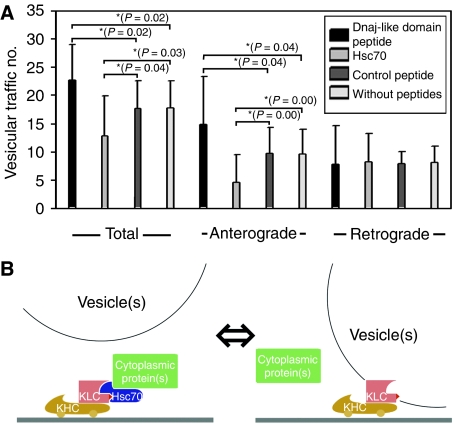
Cytoplasmic protein transport in axons ('slow axonal transport') is essential for neuronal homeostasis, and involves Kinesin-1, the same motor for membranous organelle transport ('fast axonal transport'). However, both molecular mechanisms of slow axonal transport and difference in usage of Kinesin-1 between slow and fast axonal transport have been elusive. Here, we show that slow axonal transport depends on the interaction between the DnaJ-like domain of the kinesin light chain in the Kinesin-1 motor complex and Hsc70, scaffolding between cytoplasmic proteins and Kinesin-1. The domain is within the tetratricopeptide repeat, which can bind to membranous organelles, and competitive perturbation of the domain in squid giant axons disrupted cytoplasmic protein transport and reinforced membranous organelle transport, indicating that this domain might have a function as a switchover system between slow and fast transport by Hsc70. Transgenic mice overexpressing a dominant-negative form of the domain showed delayed slow transport, accelerated fast transport and optic axonopathy. These findings provide a basis for the regulatory mechanism of intracellular transport and its intriguing implication in neuronal dysfunction.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
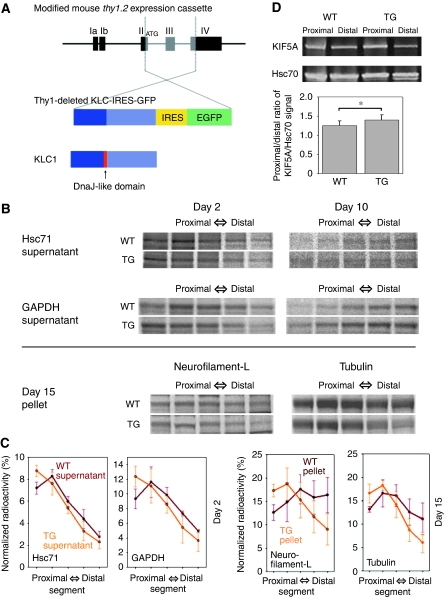
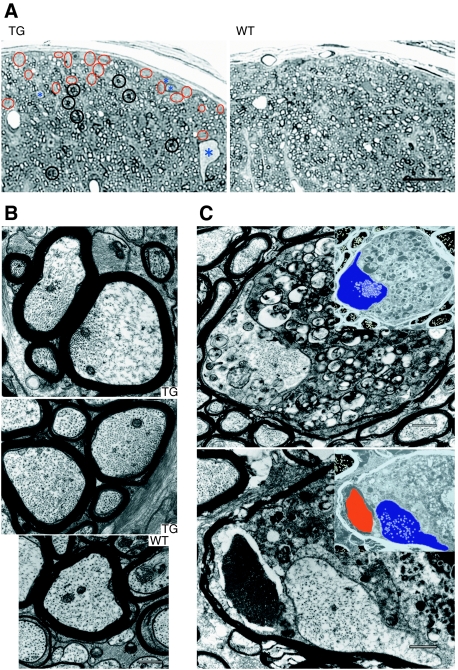
References
-
- Adachi H, Katsuno M, Minamiyama M, Sang C, Pagoulatos G, Angelidis C, Kusakabe M, Yoshiki A, Kobayashi Y, Doyu M, Sobue G (2003) Heat shock protein 70 chaperone overexpression ameliorates phenotypes of the spinal and bulbar muscular atrophy transgenic mouse model by reducing nuclear-localized mutant androgen receptor protein. J Neurosci 23: 2203–2211 - PMC - PubMed
-
- Aihara M, Lindsey JD, Weinreb RN (2002) Reduction of intraocular pressure in mouse eyes treated with latanoprost. Invest Ophthalmol Vis Sci 43: 146–150 - PubMed
-
- Andra K, Abramowski D, Duke M, Probst A, Wiederhold KH, Burki K, Goedert M, Sommer B, Staufenbiel M (1996) Expression of APP in transgenic mice: a comparison of neuron-specific promoters. Neurobiol Aging 17: 183–190 - PubMed
-
- Auluck PK, Chan HY, Trojanowski JQ, Lee VM, Bonini NM (2002) Chaperone suppression of alpha-synuclein toxicity in a Drosophila model for Parkinson's disease. Science 295: 865–868 - PubMed
-
- Baas PW, Brown A (1997) Slow axonal transport: the polymer transport model. Trends Cell Biol 7: 380–384 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
