

Distinct phases of siRNA synthesis in an endogenous RNAi pathway in C. elegans soma
- PMID: 20116306
- PMCID: PMC2838994
- DOI: 10.1016/j.molcel.2010.01.012
Distinct phases of siRNA synthesis in an endogenous RNAi pathway in C. elegans soma
Abstract
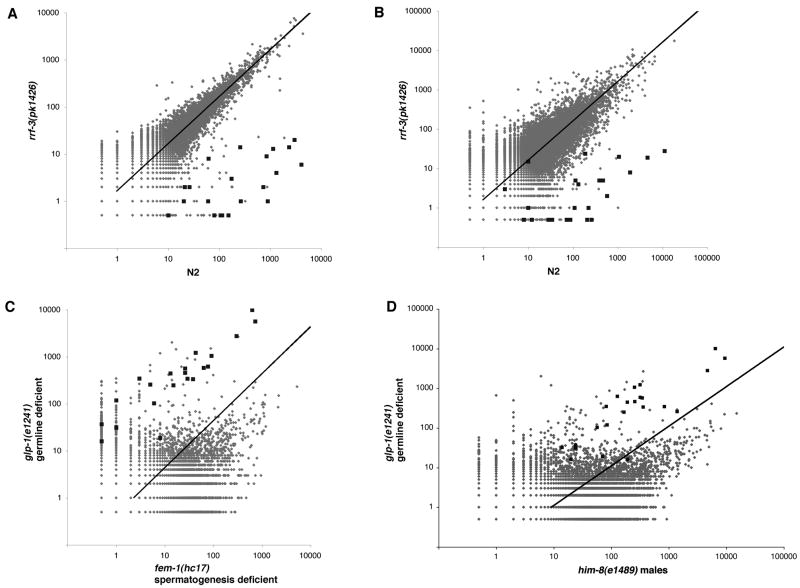
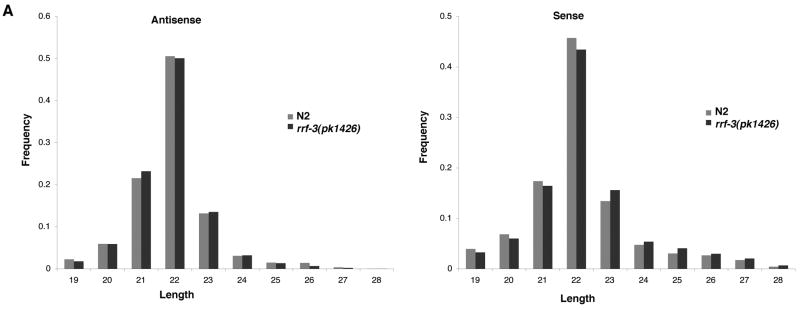
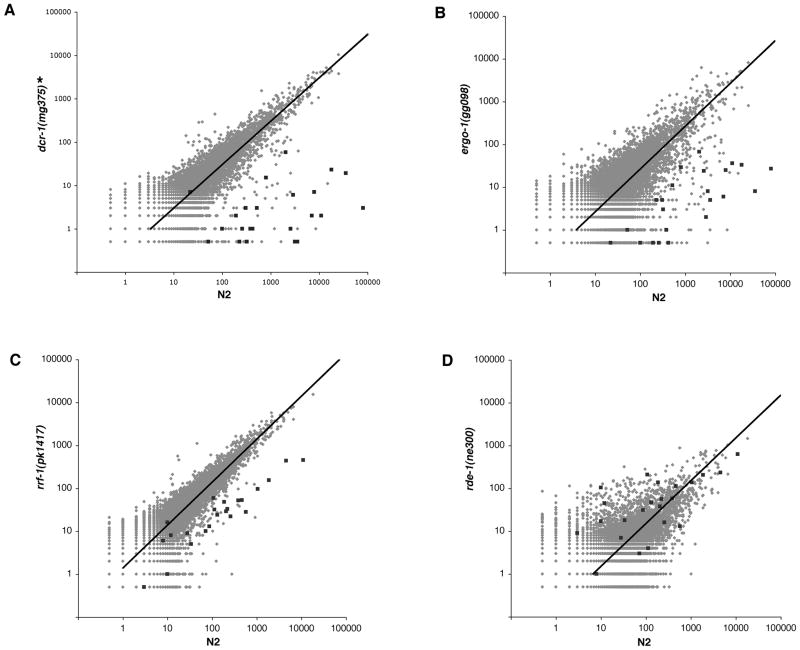
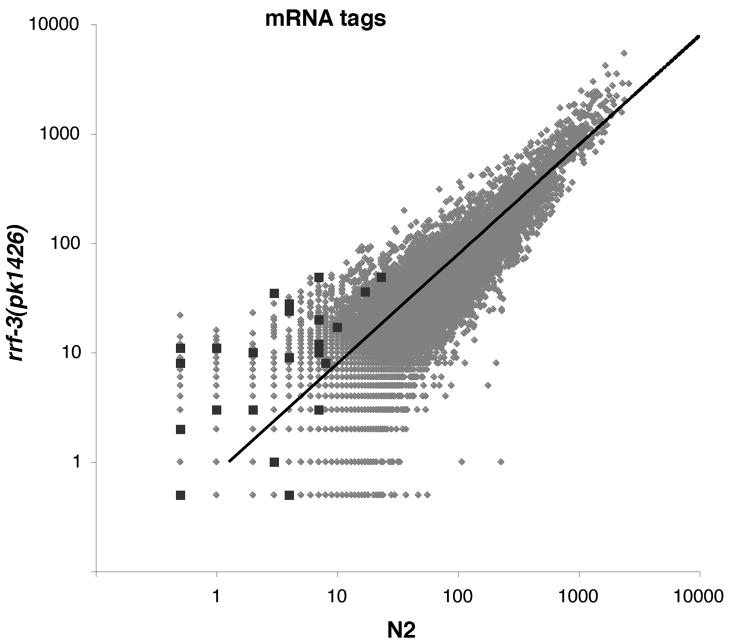
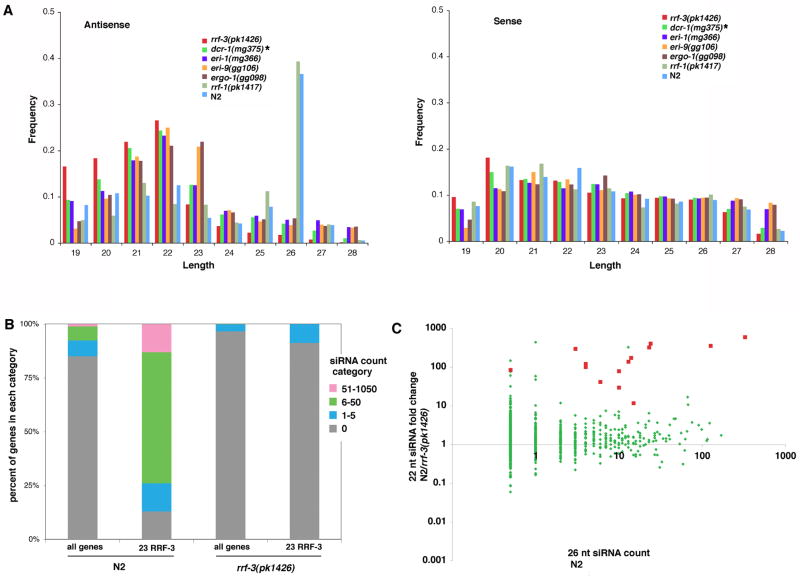
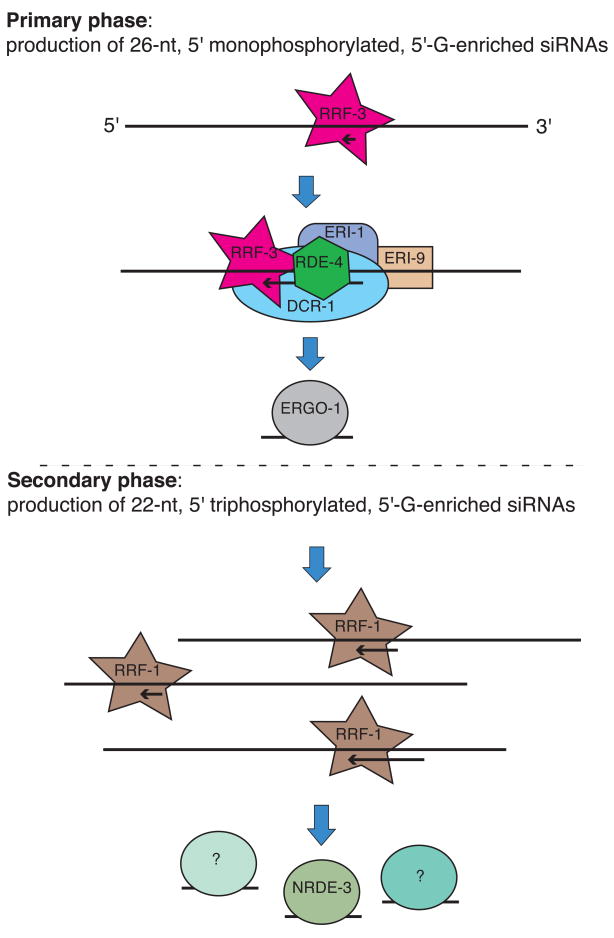
Endogenous RNA-directed RNA polymerases (RdRPs) are cellular components capable of synthesizing new complementary RNAs from existing RNA templates. We present evidence for successive engagement of two different RdRPs in an endogenous siRNA-based mechanism targeting specific mRNAs in C. elegans soma. In the initiation stage of this process, a group of mRNA species are chosen as targets for downregulation, leading to accumulation of rare 26 nt 5'-phosphorylated antisense RNAs that depend on the RdRP homolog RRF-3, the Argonaute ERGO-1, DICER, and a series of associated ("ERI") factors. This primary process leads to production of a much more abundant class of 22 nt antisense RNAs, dependent on a secondary RdRP (RRF-1) and associating with at least one distinct Argonaute (NRDE-3). The requirement for two RdRP/Argonaute combinations and initiation by a rare class of uniquely structured siRNAs in this pathway illustrate the caution and flexibility used as biological systems exploit the physiological copying of RNA.
(c) 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
An RNA-mediated silencing pathway utilizes the coordinated synthesis of two distinct populations of siRNA.Mol Cell. 2010 Mar 12;37(5):593-5. doi: 10.1016/j.molcel.2010.02.013. Mol Cell. 2010. PMID: 20227363
References
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr Biol. 2003;13:807–818. - PubMed
-
- Asikainen S, Storvik M, Lakso M, Wong G. Whole genome microarray analysis of C. elegans rrf-3 and eri-1 mutants. FEBS Lett. 2007;581:5050–5054. - PubMed
-
- Axtell MJ, Jan C, Rajagopalan R, Bartel DP. A two-hit trigger for siRNA biogenesis in plants. Cell. 2006;127:565–577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
