

Identification of stem cell transcriptional programs normally expressed in embryonic and neural stem cells in alloreactive CD8+ T cells mediating graft-versus-host disease
- PMID: 20116439
- PMCID: PMC2913321
- DOI: 10.1016/j.bbmt.2010.01.012
Identification of stem cell transcriptional programs normally expressed in embryonic and neural stem cells in alloreactive CD8+ T cells mediating graft-versus-host disease
Abstract
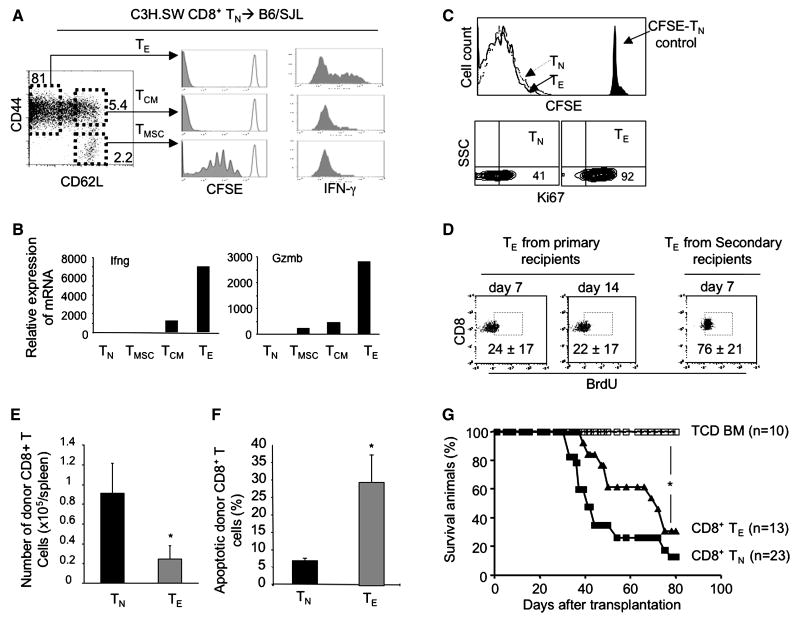
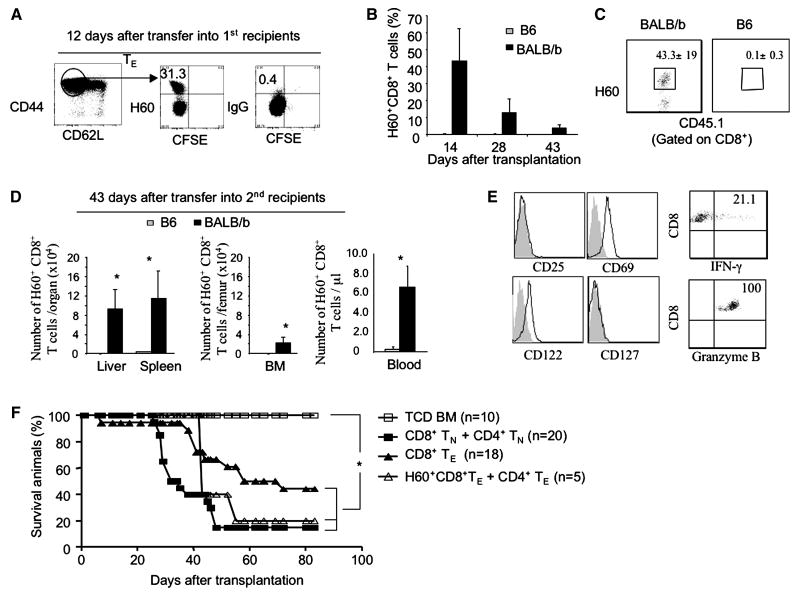
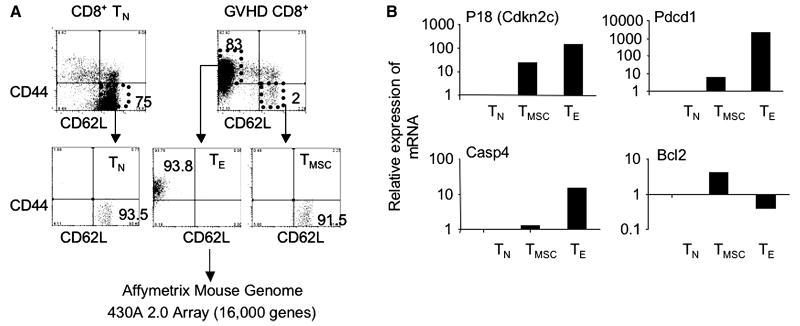
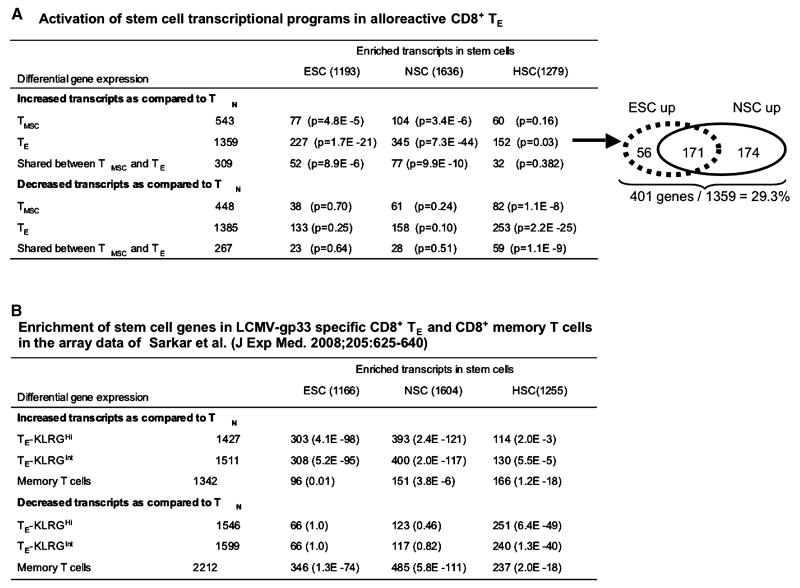
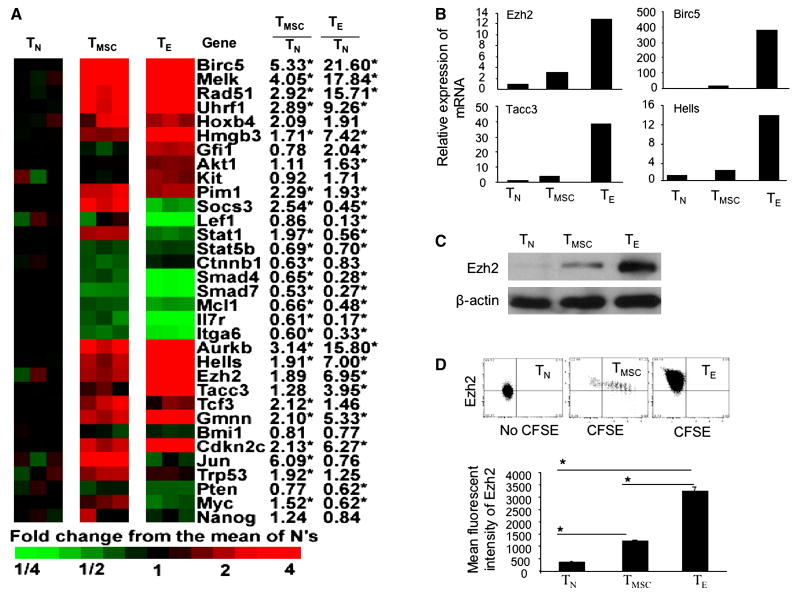
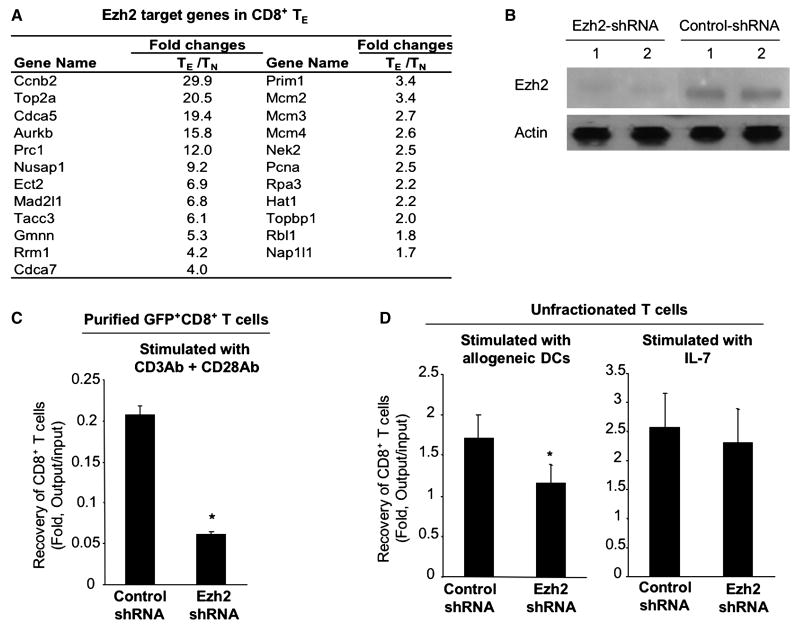
A hallmark of graft-versus-host-disease (GVHD), a life-threatening complication after allogeneic hematopoietic stem cell transplantation, is the cytopathic injury of host tissues mediated by persistent alloreactive effector T cells (T(E)). However, the mechanisms that regulate the persistence of alloreactive T(E) during GVHD remain largely unknown. Using mouse GVHD models, we demonstrate that alloreactive CD8(+) T(E) rapidly diminished in vivo when adoptively transferred into irradiated secondary congenic recipient mice. In contrast, although alloreactive CD8(+) T(E) underwent massive apoptosis upon chronic exposure to alloantigens, they proliferated in vivo in secondary allogeneic recipients, persisted, and caused severe GVHD. Thus, the continuous proliferation of alloreactive CD8(+) T(E), which is mediated by alloantigenic stimuli rather than homeostatic factors, is critical to maintaining their persistence. Gene expression profile analysis revealed that although alloreactive CD8(+) T(E) increased the expression of genes associated with cell death, they activated a group of stem cell genes normally expressed in embryonic and neural stem cells. Most of these stem cell genes are associated with cell cycle regulation, DNA replication, chromatin modification, and transcription. One of these genes, Ezh2, which encodes a chromatin modifying enzyme, was abundantly expressed in CD8(+) T(E). Silencing Ezh2 significantly reduced the proliferation of alloantigen-activated CD8(+) T cells. Thus, these findings identify that a group of stem cell genes could play important roles in sustaining terminally differentiated alloreactive CD8(+) T(E) and may be therapeutic targets for controlling GVHD.
Copyright 2010 American Society for Blood and Marrow Transplantation. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Alloreactive memory T cells are responsible for the persistence of graft-versus-host disease.J Immunol. 2005 Mar 1;174(5):3051-8. doi: 10.4049/jimmunol.174.5.3051. J Immunol. 2005. PMID: 15728519
-
Targeting of inducible costimulator (ICOS) expressed on alloreactive T cells down-regulates graft-versus-host disease (GVHD) and facilitates engraftment of allogeneic bone marrow (BM).Blood. 2005 Apr 15;105(8):3372-80. doi: 10.1182/blood-2004-10-3869. Epub 2004 Dec 23. Blood. 2005. PMID: 15618467
-
Blockade of osteopontin reduces alloreactive CD8+ T cell-mediated graft-versus-host disease.Blood. 2011 Feb 3;117(5):1723-33. doi: 10.1182/blood-2010-04-281659. Epub 2010 Nov 30. Blood. 2011. PMID: 21119110 Free PMC article.
-
Opposing effects of ICOS on graft-versus-host disease mediated by CD4 and CD8 T cells.J Immunol. 2006 Jun 15;176(12):7394-401. doi: 10.4049/jimmunol.176.12.7394. J Immunol. 2006. PMID: 16751384
-
Graft-versus-host disease impairs vaccine responses through decreased CD4+ and CD8+ T cell proliferation and increased perforin-mediated CD8+ T cell apoptosis.J Immunol. 2013 Feb 1;190(3):1351-9. doi: 10.4049/jimmunol.1200391. Epub 2012 Dec 28. J Immunol. 2013. PMID: 23275602 Free PMC article.
Cited by
-
The pleiotropic roles of EZH2 in T-cell immunity and immunotherapy.Int J Hematol. 2022 Dec;116(6):837-845. doi: 10.1007/s12185-022-03466-x. Epub 2022 Oct 21. Int J Hematol. 2022. PMID: 36271224 Review.
-
Pre- and postmortem imaging of transplanted cells.Int J Nanomedicine. 2015 Sep 2;10:5543-59. doi: 10.2147/IJN.S83557. eCollection 2015. Int J Nanomedicine. 2015. PMID: 26366076 Free PMC article. Review.
-
Inhibition of histone methylation arrests ongoing graft-versus-host disease in mice by selectively inducing apoptosis of alloreactive effector T cells.Blood. 2012 Feb 2;119(5):1274-82. doi: 10.1182/blood-2011-06-364422. Epub 2011 Nov 23. Blood. 2012. PMID: 22117046 Free PMC article.
-
T-Bet and Eomes Regulate the Balance between the Effector/Central Memory T Cells versus Memory Stem Like T Cells.PLoS One. 2013 Jun 27;8(6):e67401. doi: 10.1371/journal.pone.0067401. Print 2013. PLoS One. 2013. PMID: 23826287 Free PMC article.
-
The molecular basis of the memory T cell response: differential gene expression and its epigenetic regulation.Nat Rev Immunol. 2012 Mar 16;12(4):306-15. doi: 10.1038/nri3173. Nat Rev Immunol. 2012. PMID: 22421787 Free PMC article. Review.
References
-
- Fearon DT, Manders P, Wagner SD. Arrested differentiation, the self-renewing memory lymphocyte, and vaccination. Science. 2001;293:248–250. - PubMed
-
- Kaech SM, Hemby S, Kersh E, Ahmed R. Molecular and functional profiling of memory CD8 T cell differentiation. Cell. 2002;111:837–851. - PubMed
-
- Joshi NS, Kaech SM. Effector CD8 T cell development: a balancing act between memory cell potential and terminal differentiation. J Immunol. 2008;180:1309–1315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
