

TRAUCO, a Trithorax-group gene homologue, is required for early embryogenesis in Arabidopsis thaliana
- PMID: 20118203
- PMCID: PMC2826662
- DOI: 10.1093/jxb/erp396
TRAUCO, a Trithorax-group gene homologue, is required for early embryogenesis in Arabidopsis thaliana
Abstract
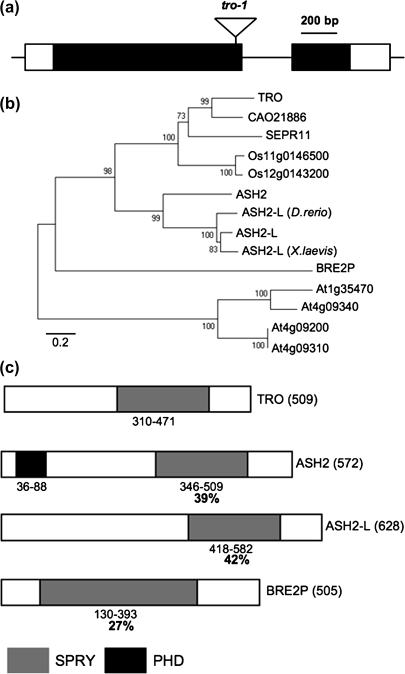
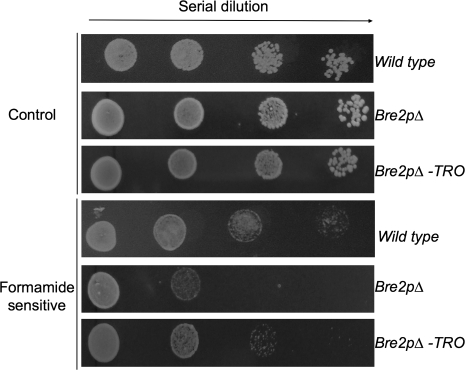
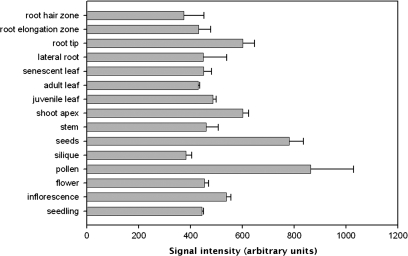
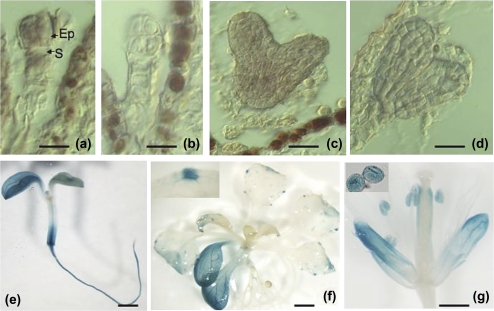
Embryogenesis is a critical stage during the plant life cycle in which a unicellular zygote develops into a multicellular organism. Co-ordinated gene expression is thus necessary for proper embryo development. Polycomb and Trithorax group genes are members of evolutionarily conserved machinery that maintains the correct expression patterns of key developmental regulators by repressing and activating gene transcription. TRAUCO (TRO), a gene homologous to the Trithorax group of genes that can functionally complement a BRE2P yeast mutant, has been identified in Arabidopsis thaliana. It is demonstrated that TRO is a nuclear gene product expressed during embryogenesis, and loss of TRO function leads to impaired early embryo development. Embryos that arrested at the globular stage in the tro-1 mutant allele were fully rescued by a TRO expression clone, a demonstration that the tro-1 mutation is a true loss-of-function in TRO. Our data have established that TRO is the first trithorax-group gene homologue in plants that is required for early embryogenesis.
Figures
Similar articles
-
Arabidopsis thaliana GEX1 has dual functions in gametophyte development and early embryogenesis.Plant J. 2011 Nov;68(4):620-32. doi: 10.1111/j.1365-313X.2011.04713.x. Epub 2011 Aug 30. Plant J. 2011. PMID: 21831199
-
Cell-by-cell developmental transition from embryo to post-germination phase revealed by heterochronic gene expression and ER-body formation in Arabidopsis leafy cotyledon mutants.Plant Cell Physiol. 2014 Dec;55(12):2112-25. doi: 10.1093/pcp/pcu139. Epub 2014 Oct 4. Plant Cell Physiol. 2014. PMID: 25282558
-
The Arabidopsis NF-YA3 and NF-YA8 genes are functionally redundant and are required in early embryogenesis.PLoS One. 2013 Nov 26;8(11):e82043. doi: 10.1371/journal.pone.0082043. eCollection 2013. PLoS One. 2013. PMID: 24303077 Free PMC article.
-
Polycomb group and trithorax group proteins in Arabidopsis.Biochim Biophys Acta. 2007 May-Jun;1769(5-6):375-82. doi: 10.1016/j.bbaexp.2007.01.010. Epub 2007 Feb 7. Biochim Biophys Acta. 2007. PMID: 17363079 Review.
-
The origin of the plant body axis.Curr Opin Plant Biol. 2012 Dec;15(6):578-84. doi: 10.1016/j.pbi.2012.08.001. Epub 2012 Aug 22. Curr Opin Plant Biol. 2012. PMID: 22921364 Review.
Cited by
-
Arabidopsis S2Lb links AtCOMPASS-like and SDG2 activity in H3K4me3 independently from histone H2B monoubiquitination.Genome Biol. 2019 May 21;20(1):100. doi: 10.1186/s13059-019-1705-4. Genome Biol. 2019. PMID: 31113491 Free PMC article.
-
HAG1 and SWI3A/B control of male germ line development in P. patens suggests conservation of epigenetic reproductive control across land plants.Plant Reprod. 2021 Jun;34(2):149-173. doi: 10.1007/s00497-021-00409-0. Epub 2021 Apr 11. Plant Reprod. 2021. PMID: 33839924 Free PMC article.
-
Arabidopsis COMPASS-like complexes mediate histone H3 lysine-4 trimethylation to control floral transition and plant development.PLoS Genet. 2011 Mar;7(3):e1001330. doi: 10.1371/journal.pgen.1001330. Epub 2011 Mar 10. PLoS Genet. 2011. PMID: 21423667 Free PMC article.
-
Comprehensive analysis of Alfin-like transcription factors associated with drought and salt stresses in wheat (Triticum aestivum L.).BMC Genomics. 2024 Jul 17;25(1):701. doi: 10.1186/s12864-024-10557-y. BMC Genomics. 2024. PMID: 39020295 Free PMC article.
References
-
- Alexander MP. Differential staining of aborted and non-aborted pollen. Stain Technology. 1969;44:117–122. - PubMed
-
- Alvarez-Venegas R, Pien S, Sadder M, Witmer X, Grossniklaus U, Avramova Z. ATX-1, an Arabidopsis homolog of trithorax, activates flower homeotic genes. Current Biology. 2003;13:627–637. - PubMed
-
- Aquea F, Arce-Johnson P. Identification of genes expressed during early somatic embryogenesis in Pinus radiata. Plant Physiology and Biochemistry. 2008;46:559–568. - PubMed
-
- Aquea F, Matte JP, Gutiérrez F, Rico S, Lamprecht M, Sánchez C, Arce-Johnson P. Molecular characterization of a Trithorax-group homologue gene from Pinus radiata. Plant Cell Reporter. 2009;28:1531–1538. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
