

Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin
- PMID: 20118940
- PMCID: PMC2831214
- DOI: 10.1038/nchembio.304
Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin
Abstract
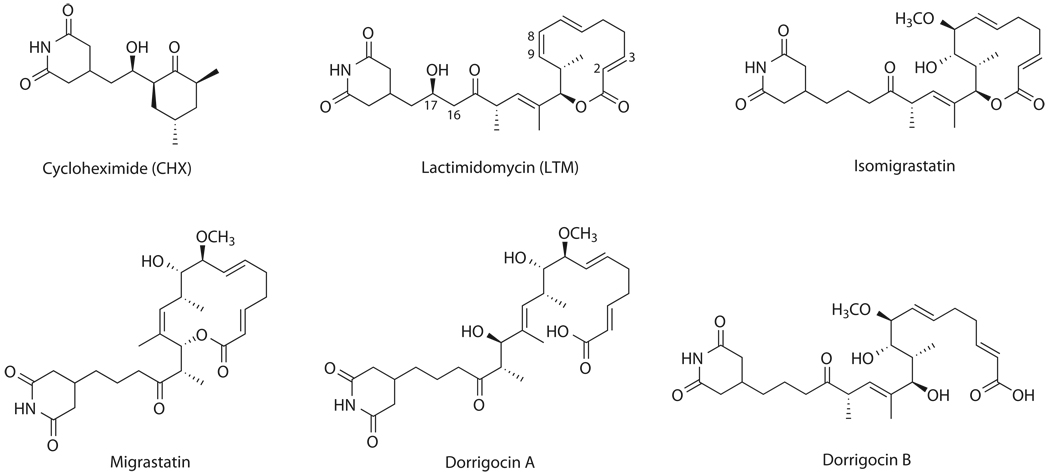
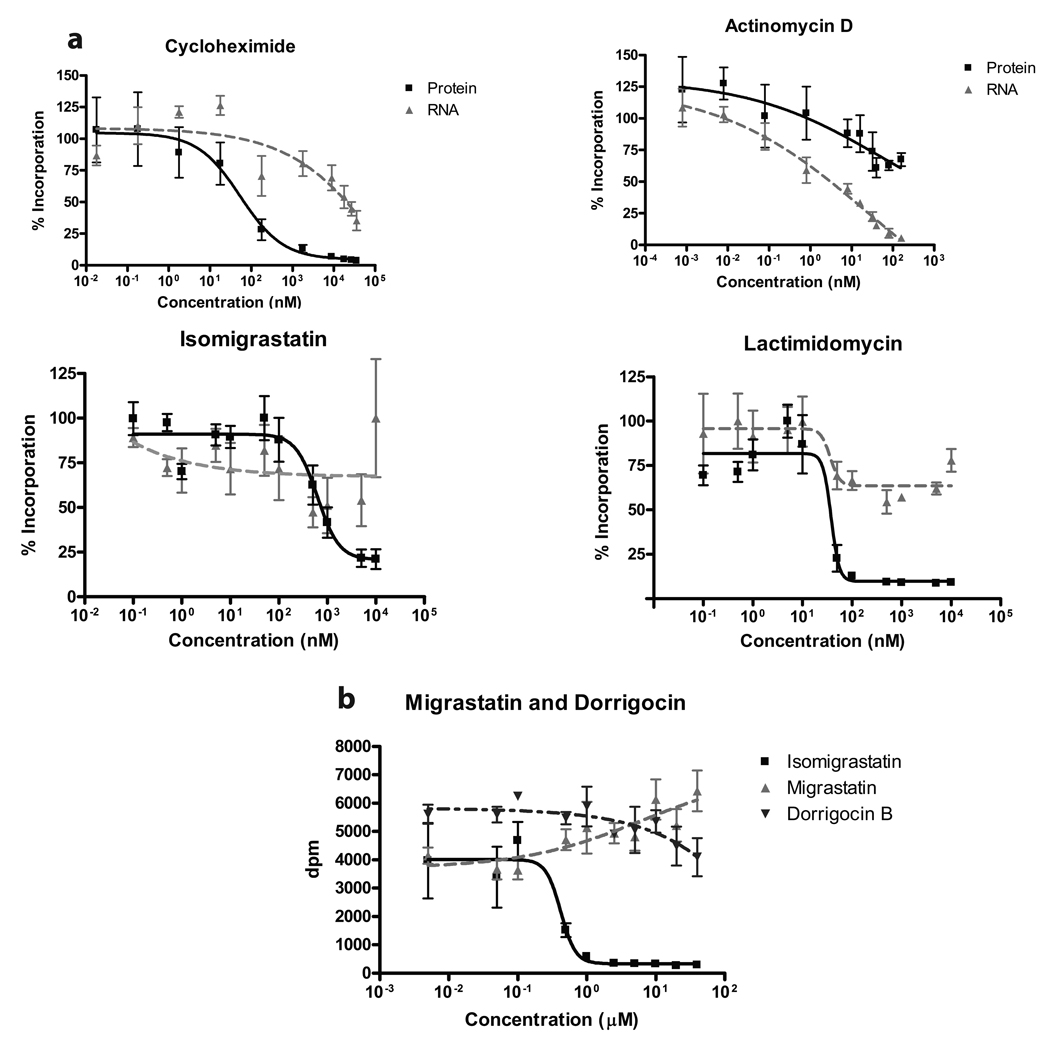
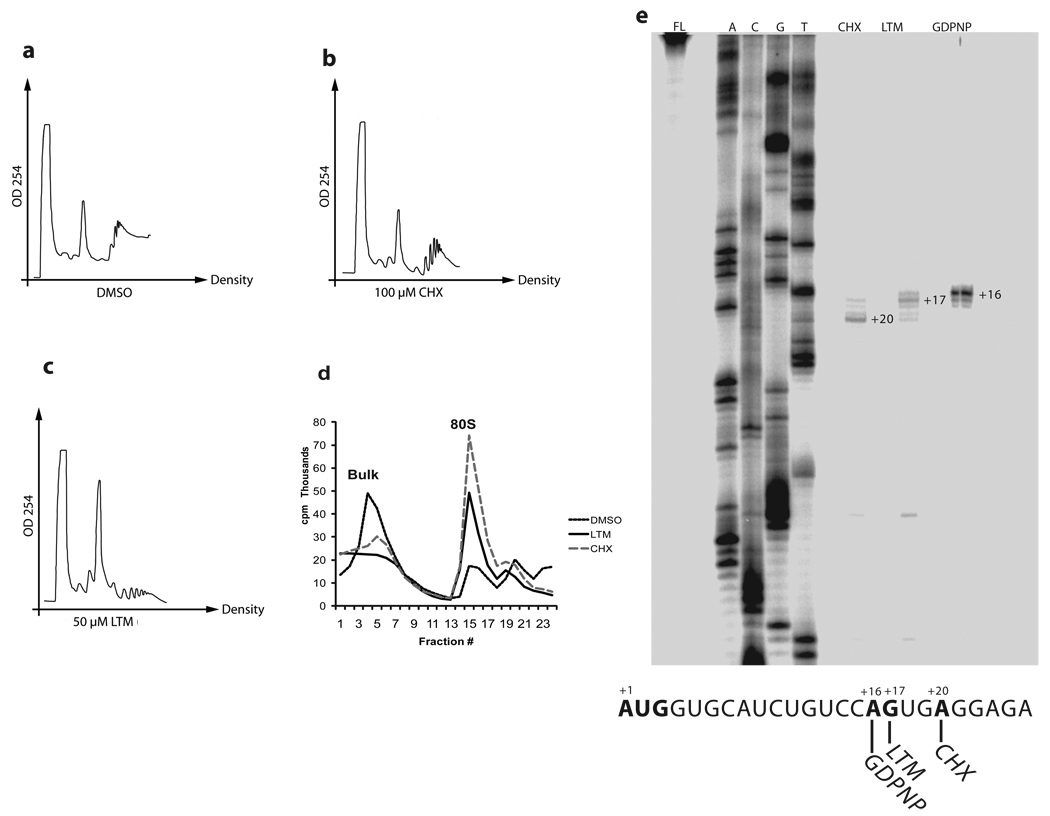
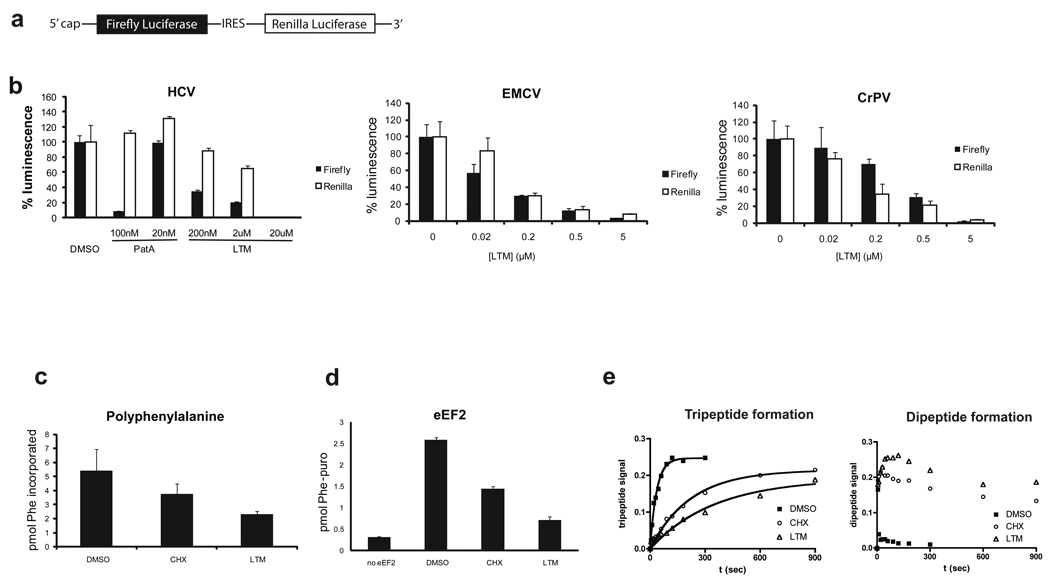
Although the protein synthesis inhibitor cycloheximide (CHX) has been known for decades, its precise mechanism of action remains incompletely understood. The glutarimide portion of CHX is seen in a family of structurally related natural products including migrastatin, isomigrastatin and lactimidomycin (LTM). We found that LTM, isomigrastatin and analogs have a potent antiproliferative effect on tumor cell lines and selectively inhibit translation. A systematic comparative study of the effects of CHX and LTM on protein synthesis revealed both similarities and differences between the two inhibitors. Both LTM and CHX were found to block the translocation step in elongation. Footprinting experiments revealed protection of a single cytidine nucleotide (C3993) in the E-site of the 60S ribosomal subunit, thus defining a common binding pocket for the two inhibitors in the ribosome. These results shed new light on the molecular mechanism of inhibition of translation elongation by both CHX and LTM.
Figures
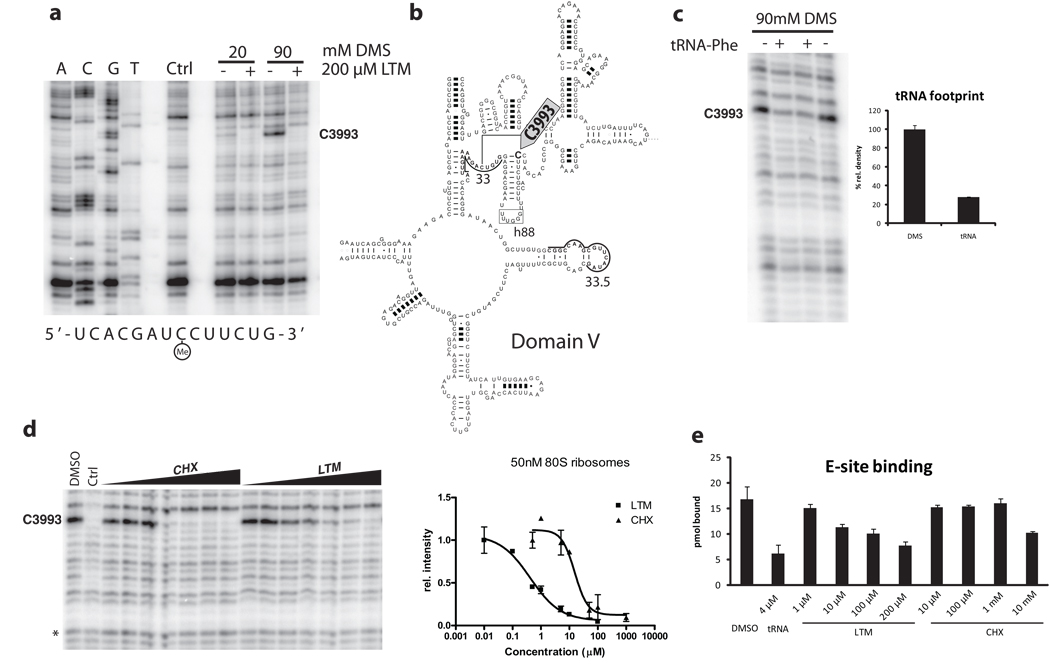
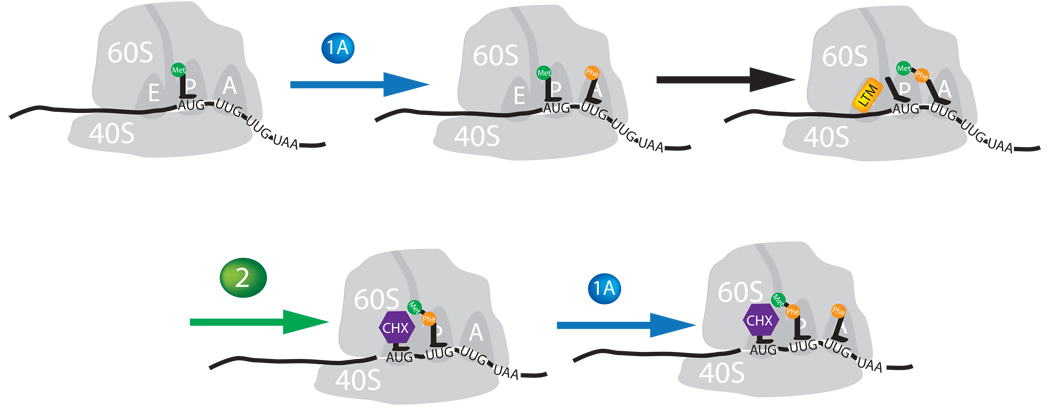
References
-
- Poehlsgaard J, Douthwaite S. The bacterial ribosome as a target for antibiotics. Nat Rev Microbiol. 2005;3:870–881. - PubMed
-
- Obrig TG, Culp WJ, McKeehan WL, Hardesty B. The mechanism by which cycloheximide and related glutarimide antibiotics inhibit peptide synthesis on reticulocyte ribosomes. J Biol Chem. 1971;246:174–181. - PubMed
-
- Ju J, Lim SK, Jiang H, Seo JW, Shen B. Iso-migrastatin congeners from Streptomyces platensis and generation of a glutarimide polyketide library featuring the dorrigocin, lactimidomycin, migrastatin, and NK30424 scaffolds. J Am Chem Soc. 2005;127:11930–11931. - PubMed
-
- Sugawara K, et al. Lactimidomycin, a new glutarimide group antibiotic. Production, isolation, structure and biological activity. J Antibiot (Tokyo) 1992;45:1433–1441. - PubMed
Associated data
- PubChem-Substance/87226503
- PubChem-Substance/87226504
- PubChem-Substance/87226505
- PubChem-Substance/87226506
- PubChem-Substance/87226507
- PubChem-Substance/87226508
- PubChem-Substance/87226509
- PubChem-Substance/87226510
- PubChem-Substance/87226511
- PubChem-Substance/87226512
- PubChem-Substance/87226513
- PubChem-Substance/87226514
- PubChem-Substance/87226515
- PubChem-Substance/87226516
- PubChem-Substance/87226517
- PubChem-Substance/87226518
- PubChem-Substance/87226519
- PubChem-Substance/87226520
- PubChem-Substance/87226521
- PubChem-Substance/87226522
- PubChem-Substance/87226523
- PubChem-Substance/87226524
- PubChem-Substance/87226537
- PubChem-Substance/87226538
- PubChem-Substance/87226525
- PubChem-Substance/87226526
- PubChem-Substance/87226527
- PubChem-Substance/87226528
- PubChem-Substance/87226529
- PubChem-Substance/87226530
- PubChem-Substance/87226531
- PubChem-Substance/87226532
- PubChem-Substance/87226533
- PubChem-Substance/87226534
- PubChem-Substance/87226535
- PubChem-Substance/87226536
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
