

Alpha-2 noradrenergic receptor activation inhibits the hyperpolarization-activated cation current (Ih) in neurons of the ventral tegmental area
- PMID: 20122999
- PMCID: PMC2854519
- DOI: 10.1016/j.neuroscience.2010.01.052
Alpha-2 noradrenergic receptor activation inhibits the hyperpolarization-activated cation current (Ih) in neurons of the ventral tegmental area
Abstract
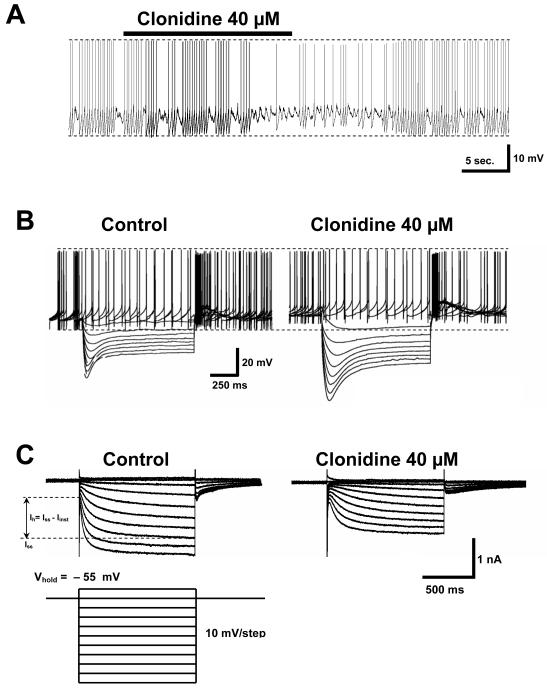
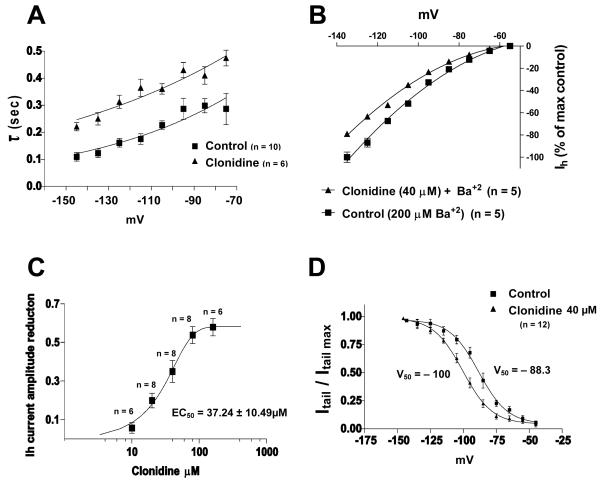
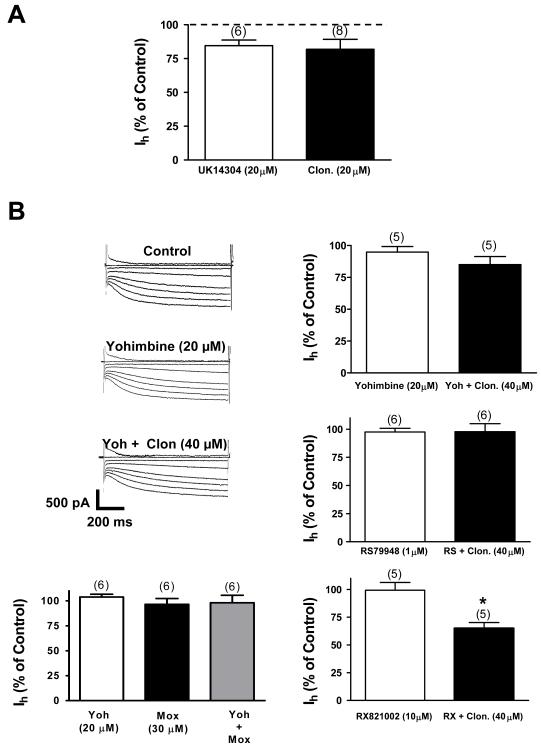
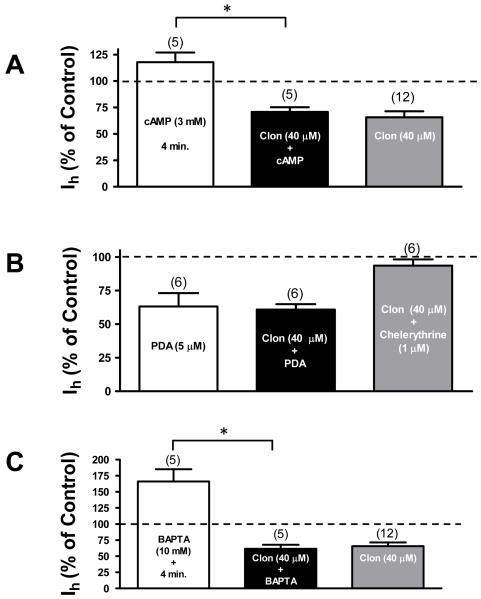
The ventral tegmental area (VTA) is the source of dopaminergic projections innervating cortical structures and ventral forebrain. Dysfunction of this mesocorticolimbic system is critically involved in psychiatric disorders such as addiction and schizophrenia. Changes in VTA dopamine (DA) neuronal activity can alter neurotransmitter release at target regions which modify information processing in the reward circuit. Here we studied the effect of alpha-2 noradrenergic receptor activation on the hyperpolarization-activated cation current (I(h)) in DA neurons of the rat VTA. Brain slice preparations using whole-cell current and voltage-clamp techniques were employed. Clonidine and UK14304 (alpha-2 receptor selective agonists) were found to decrease I(h) amplitude and to slow its rate of activation indicating a negative shift in the current's voltage dependence. Two non-subtype-selective alpha-2 receptor antagonists, yohimbine and RS79948, prevented the effects of alpha-2 receptor activation. RX821002, a noradrenergic antagonist specific for alpha-2A and alpha-2D did not prevent I(h) inhibition. This result suggests that clonidine might be acting via an alpha-2C subtype since this receptor is the most abundant variant in the VTA. Analysis of a second messenger system associated with the alpha-2 receptor revealed that I(h) inhibition is independent of cyclic AMP (cAMP) and resulted from the activation of protein kinase C. It is suggested that the alpha-2 mediated hyperpolarizing shift in I(h) voltage dependence can facilitate the transition from pacemaker firing to afferent-driven burst activity. This transition may play a key role on the changes in synaptic plasticity that occurs in the mesocorticolimbic system under pathological conditions.
(c) 2010 IBRO. Published by Elsevier Ltd. All rights reserved.
Figures
References
-
- Aghajanian G, Bunney BC. In: Usdin E, Snyder SH, editors. Central dopaminergic neurons: Neurophysiologic identification and responses to drugs; Frontiers in catecholamine research; proceedings; Elmsford, N.Y., Pergamon Press. 1973.pp. 643–648.
-
- Berger T, Senn W, Luscher RL. Hyperpolarization-activated current Ih disconnects somatic and dendritic spike initiation zones in layer V pyramidal neurons. J. Neurophysiol. 2003;90:2428–2437. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
