

Differential roles of regulatory light chain and myosin binding protein-C phosphorylations in the modulation of cardiac force development
- PMID: 20123786
- PMCID: PMC2849963
- DOI: 10.1113/jphysiol.2009.183897
Differential roles of regulatory light chain and myosin binding protein-C phosphorylations in the modulation of cardiac force development
Abstract
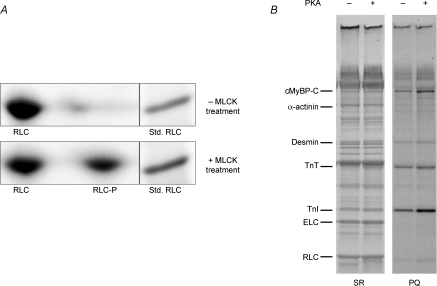
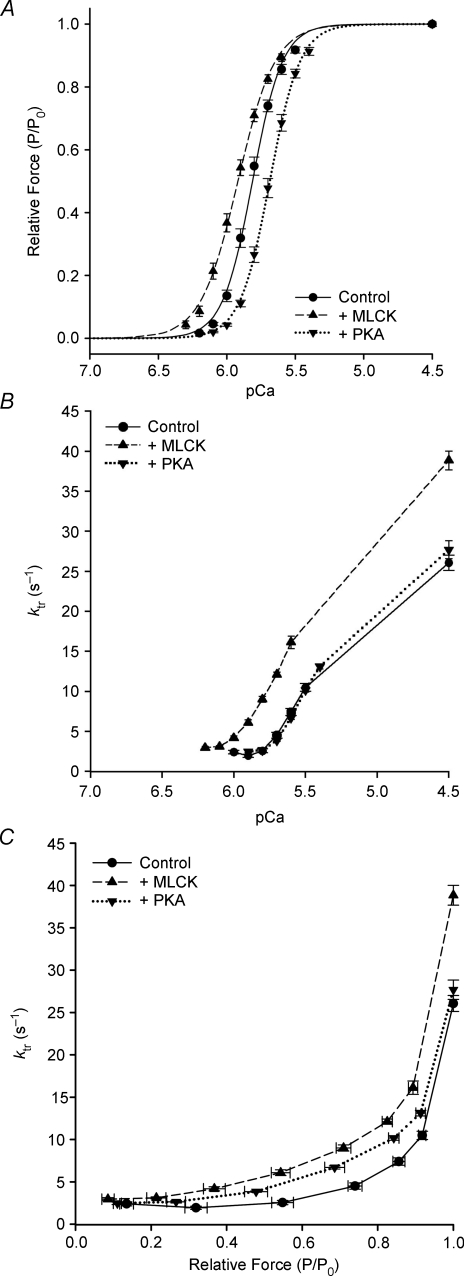
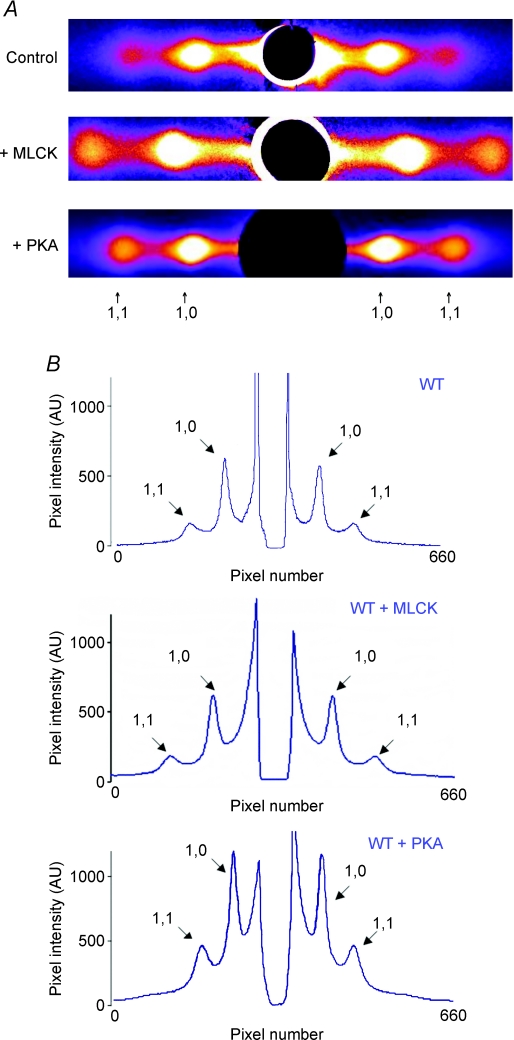
Phosphorylation of myosin regulatory light chain (RLC) by myosin light chain kinase (MLCK) and myosin binding protein-C (cMyBP-C) by protein kinase A (PKA) independently accelerate the kinetics of force development in ventricular myocardium. However, while MLCK treatment has been shown to increase the Ca(2+) sensitivity of force (pCa(50)), PKA treatment has been shown to decrease pCa(50), presumably due to cardiac troponin I phosphorylation. Further, MLCK treatment increases Ca(2+)-independent force and maximum Ca(2+)-activated force, whereas PKA treatment has no effect on either force. To investigate the structural basis underlying the kinase-specific differential effects on steady-state force, we used synchrotron low-angle X-ray diffraction to compare equatorial intensity ratios (I(1,1)/I(1,0)) to assess the proximity of myosin cross-bridge mass relative to actin and to compare lattice spacings (d(1,0)) to assess the inter-thick filament spacing in skinned myocardium following treatment with either MLCK or PKA. As we showed previously, PKA phosphorylation of cMyBP-C increases I(1,1)/I(1,0) and, as hypothesized, treatment with MLCK also increased I(1,1)/I(1,0), which can explain the accelerated rates of force development during activation. Importantly, interfilament spacing was reduced by 2 nm (3.5%) with MLCK treatment, but did not change with PKA treatment. Thus, RLC or cMyBP-C phosphorylation increases the proximity of cross-bridges to actin, but only RLC phosphorylation affects lattice spacing, which suggests that RLC and cMyBP-C modulate the kinetics of force development by similar structural mechanisms; however, the effect of RLC phosphorylation to increase the Ca(2+) sensitivity of force is mediated by a distinct mechanism, most probably involving changes in interfilament spacing.
Figures
References
-
- Chandra M, Dong WJ, Pan BS, Cheung HC, Solaro RJ. Effects of protein kinase A phosphorylation on signalling between cardiac troponin I and the N-terminal domain of cardiac troponin C. Biochemistry. 1997;36:13305–13311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous