

Polycomb complexes act redundantly to repress genomic repeats and genes
- PMID: 20123906
- PMCID: PMC2811828
- DOI: 10.1101/gad.544410
Polycomb complexes act redundantly to repress genomic repeats and genes
Abstract
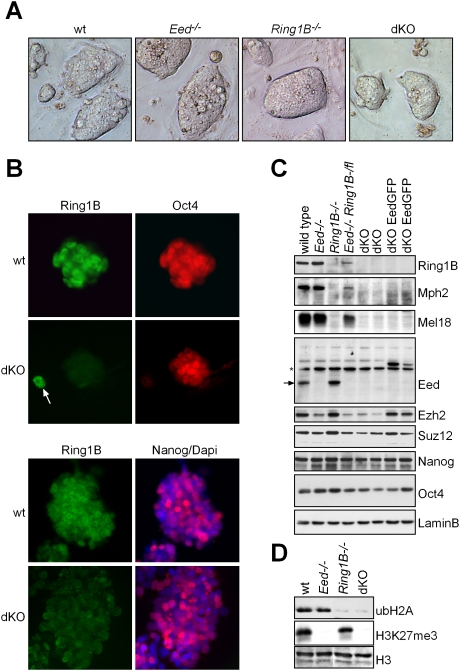
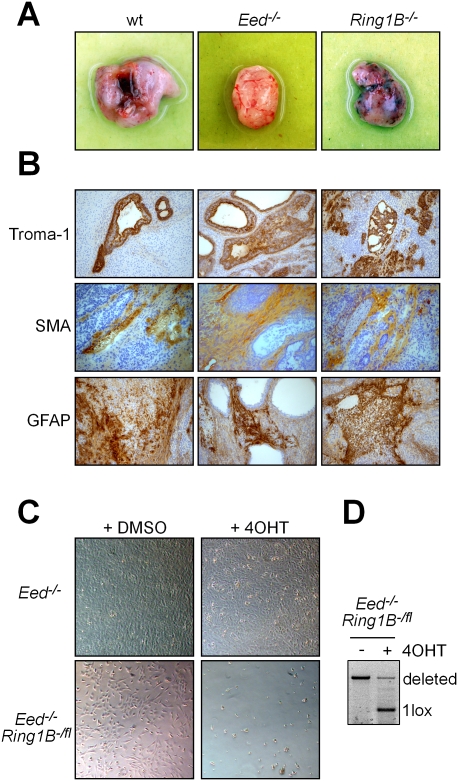
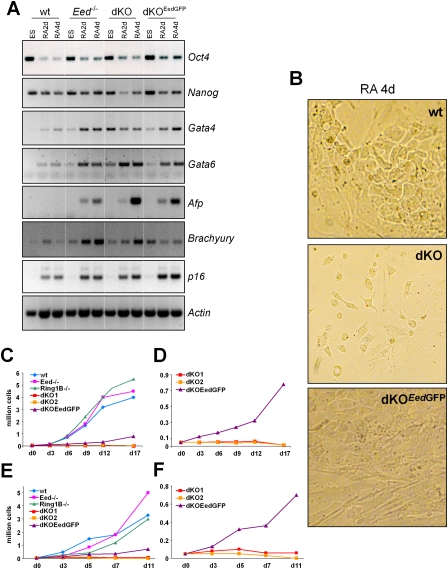
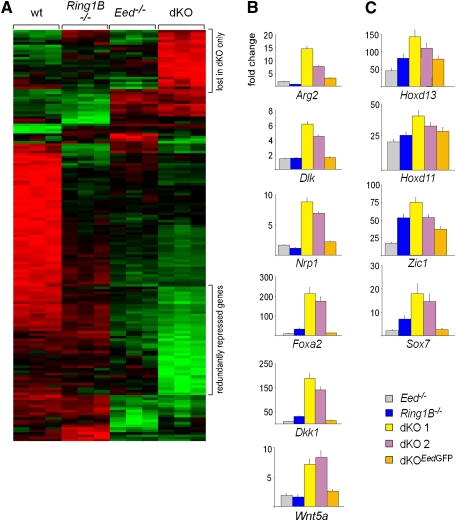
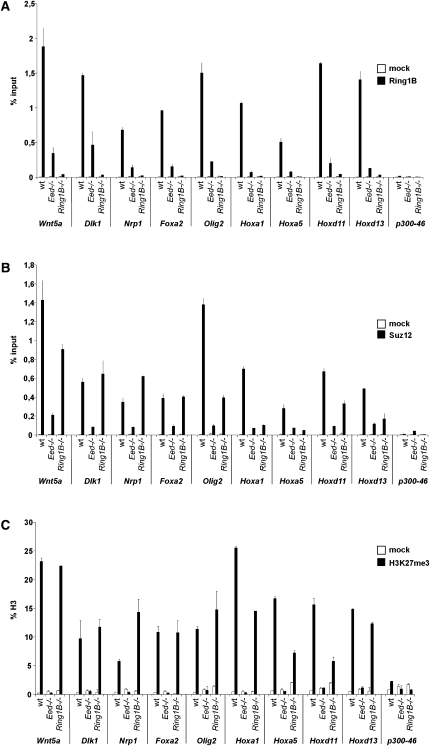
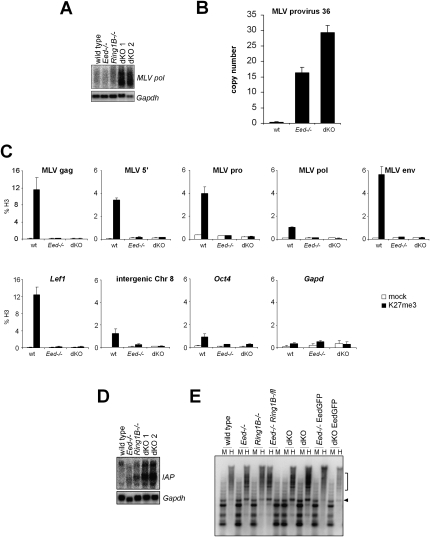
Polycomb complexes establish chromatin modifications for maintaining gene repression and are essential for embryonic development in mice. Here we use pluripotent embryonic stem (ES) cells to demonstrate an unexpected redundancy between Polycomb-repressive complex 1 (PRC1) and PRC2 during the formation of differentiated cells. ES cells lacking the function of either PRC1 or PRC2 can differentiate into cells of the three germ layers, whereas simultaneous loss of PRC1 and PRC2 abrogates differentiation. On the molecular level, the differentiation defect is caused by the derepression of a set of genes that is redundantly repressed by PRC1 and PRC2 in ES cells. Furthermore, we find that genomic repeats are Polycomb targets and show that, in the absence of Polycomb complexes, endogenous murine leukemia virus elements can mobilize. This indicates a contribution of the Polycomb group system to the defense against parasitic DNA, and a potential role of genomic repeats in Polycomb-mediated gene regulation.
Figures
References
-
- Arrigoni R, Alam SL, Wamstad JA, Bardwell VJ, Sundquist WI, Schreiber-Agus N. The Polycomb-associated protein Rybp is a ubiquitin binding protein. FEBS Lett. 2006;580:6233–6241. - PubMed
-
- Atsuta T, Fujimura S, Moriya H, Vidal M, Akasaka T, Koseki H. Production of monoclonal antibodies against mammalian Ring1B proteins. Hybridoma. 2001;20:43–46. - PubMed
-
- Boyer LA, Mathur D, Jaenisch R. Molecular control of pluripotency. Curr Opin Genet Dev. 2006a;16:455–462. - PubMed
-
- Boyer LA, Plath K, Zeitlinger J, Brambrink T, Medeiros LA, Lee TI, Levine SS, Wernig M, Tajonar A, Ray MK, et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature. 2006b;441:349–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases