

Regulation of Drosophila vasa in vivo through paralogous cullin-RING E3 ligase specificity receptors
- PMID: 20123973
- PMCID: PMC2838069
- DOI: 10.1128/MCB.01100-09
Regulation of Drosophila vasa in vivo through paralogous cullin-RING E3 ligase specificity receptors
Abstract
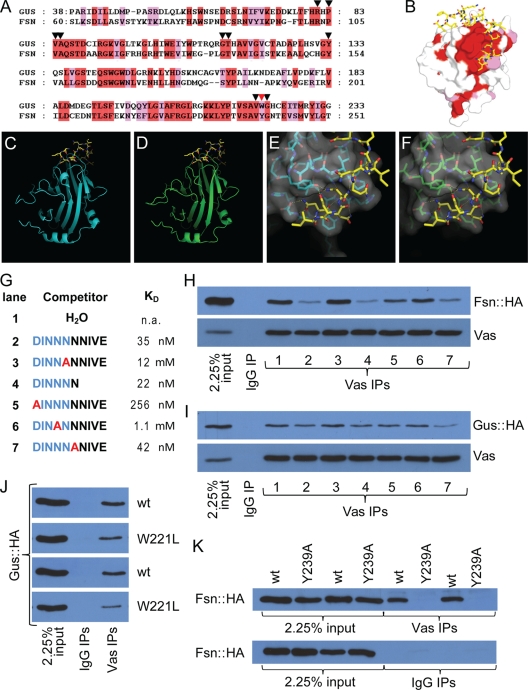
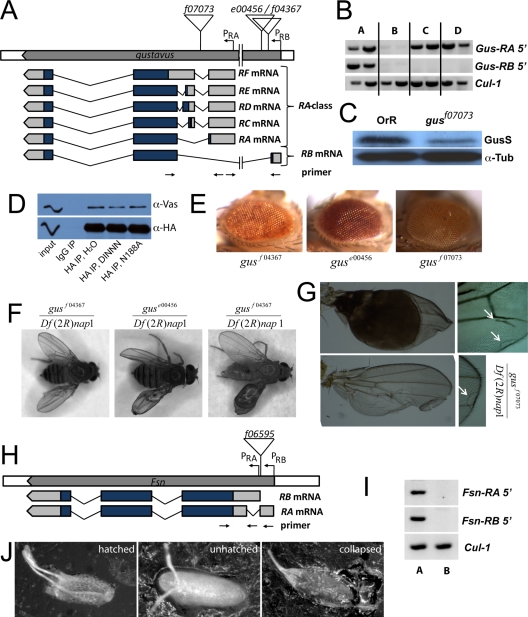
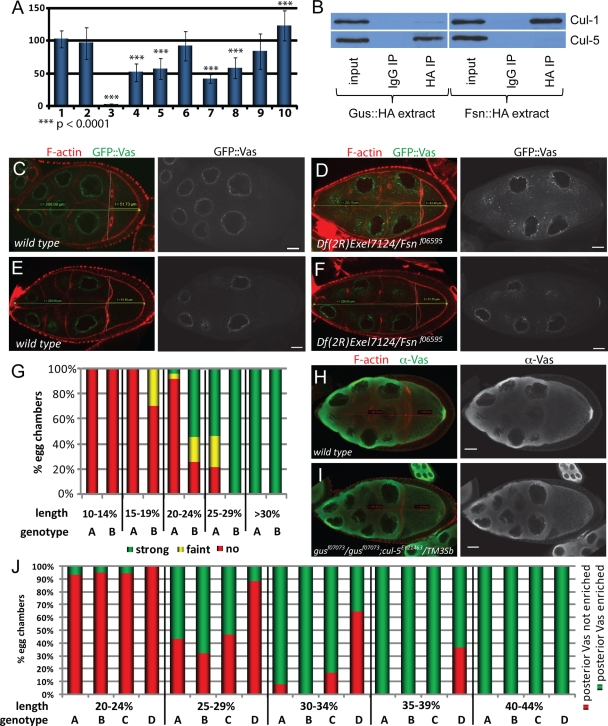
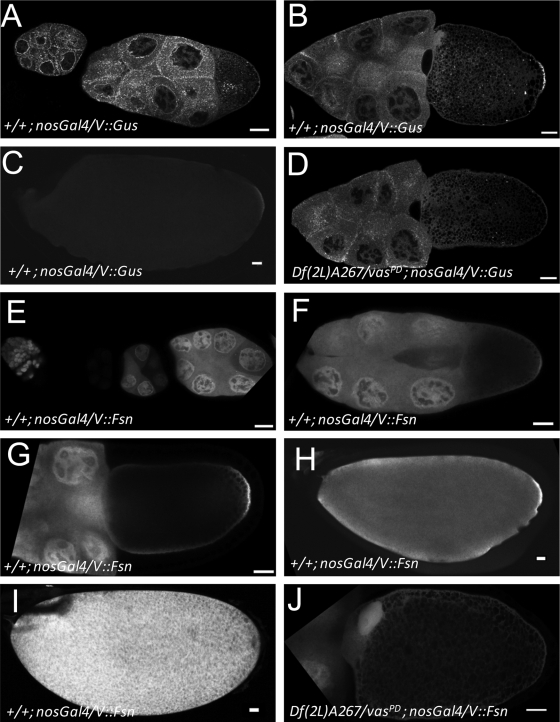
In Drosophila species, molecular asymmetries guiding embryonic development are established maternally. Vasa, a DEAD-box RNA helicase, accumulates in the posterior pole plasm, where it is required for embryonic germ cell specification. Maintenance of Vasa at the posterior pole requires the deubiquitinating enzyme Fat facets, which protects Vasa from degradation. Here, we found that Gustavus (Gus) and Fsn, two ubiquitin Cullin-RING E3 ligase specificity receptors, bind to the same motif on Vasa through their paralogous B30.2/SPRY domains. Both Gus and Fsn accumulate in the pole plasm in a Vasa-dependent manner. Posterior Vasa accumulation is precocious in Fsn mutant oocytes; Fsn overexpression reduces ovarian Vasa levels, and embryos from Fsn-overexpressing females form fewer primordial germ cells (PGCs); thus, Fsn destabilizes Vasa. In contrast, endogenous Gus may promote Vasa activity in the pole plasm, as gus females produce embryos with fewer PGCs, and posterior accumulation of Vas is delayed in gus mutant oocytes that also lack one copy of cullin-5. We propose that Fsn- and Gus-containing E3 ligase complexes contribute to establishing a fine-tuned steady state of Vasa ubiquitination that influences the kinetics of posterior Vasa deployment.
Figures
Similar articles
-
VASA localization requires the SPRY-domain and SOCS-box containing protein, GUSTAVUS.Dev Cell. 2002 Dec;3(6):865-76. doi: 10.1016/s1534-5807(02)00361-1. Dev Cell. 2002. PMID: 12479811
-
Identifying determinants of cullin binding specificity among the three functionally different Drosophila melanogaster Roc proteins via domain swapping.PLoS One. 2008 Aug 13;3(8):e2918. doi: 10.1371/journal.pone.0002918. PLoS One. 2008. PMID: 18698375 Free PMC article.
-
Fat facets interacts with vasa in the Drosophila pole plasm and protects it from degradation.Curr Biol. 2003 Oct 28;13(21):1905-9. doi: 10.1016/j.cub.2003.10.026. Curr Biol. 2003. PMID: 14588248
-
Multiple Functions of the DEAD-Box Helicase Vasa in Drosophila Oogenesis.Results Probl Cell Differ. 2017;63:127-147. doi: 10.1007/978-3-319-60855-6_6. Results Probl Cell Differ. 2017. PMID: 28779316 Review.
-
The DEAD-box helicase Vasa: evidence for a multiplicity of functions in RNA processes and developmental biology.Biochim Biophys Acta. 2013 Aug;1829(8):810-6. doi: 10.1016/j.bbagrm.2013.04.005. Epub 2013 Apr 13. Biochim Biophys Acta. 2013. PMID: 23587717 Review.
Cited by
-
Protein degradation machinery is present broadly during early development in the sea urchin.Gene Expr Patterns. 2014 Jul;15(2):135-41. doi: 10.1016/j.gep.2014.06.002. Epub 2014 Jun 22. Gene Expr Patterns. 2014. PMID: 24963879 Free PMC article.
-
C-terminal residues specific to Vasa among DEAD-box helicases are required for its functions in piRNA biogenesis and embryonic patterning.Dev Genes Evol. 2016 Nov;226(6):401-412. doi: 10.1007/s00427-016-0560-5. Epub 2016 Aug 29. Dev Genes Evol. 2016. PMID: 27572922
-
Post-translational regulation by gustavus contributes to selective Vasa protein accumulation in multipotent cells during embryogenesis.Dev Biol. 2011 Jan 15;349(2):440-50. doi: 10.1016/j.ydbio.2010.10.031. Epub 2010 Oct 28. Dev Biol. 2011. PMID: 21035437 Free PMC article.
-
SPSB1, a Novel Negative Regulator of the Transforming Growth Factor-β Signaling Pathway Targeting the Type II Receptor.J Biol Chem. 2015 Jul 17;290(29):17894-17908. doi: 10.1074/jbc.M114.607184. Epub 2015 Jun 1. J Biol Chem. 2015. PMID: 26032413 Free PMC article.
-
Structure and function of the SPRY/B30.2 domain proteins involved in innate immunity.Protein Sci. 2013 Jan;22(1):1-10. doi: 10.1002/pro.2185. Protein Sci. 2013. PMID: 23139046 Free PMC article. Review.
References
-
- Besse, F., and A. Ephrussi. 2008. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9:971-980. - PubMed
-
- Carrera, P., O. Johnstone, A. Nakamura, J. Casanova, H. Jackle, and P. Lasko. 2000. VASA mediates translation through interaction with a Drosophila yIF2 homolog. Mol. Cell 5:181-187. - PubMed
-
- Doronkin, S., I. Djagaeva, and S. K. Beckendorf. 2002. CSN5/Jab1 mutations affect axis formation in the Drosophila oocyte by activating a meiotic checkpoint. Development 129:5053-5064. - PubMed
-
- Gao, M. X., E. H. Liao, B. Yu, Y. Wang, M. Zhen, and W. B. Derry. 2008. The SCF FSN-1 ubiquitin ligase controls germline apoptosis through CEP-1/p53 in C. elegans. Cell Death Differ. 15:1054-1062. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases