

Proteus mirabilis pmrI, an RppA-regulated gene necessary for polymyxin B resistance, biofilm formation, and urothelial cell invasion
- PMID: 20123999
- PMCID: PMC2849355
- DOI: 10.1128/AAC.01219-09
Proteus mirabilis pmrI, an RppA-regulated gene necessary for polymyxin B resistance, biofilm formation, and urothelial cell invasion
Abstract
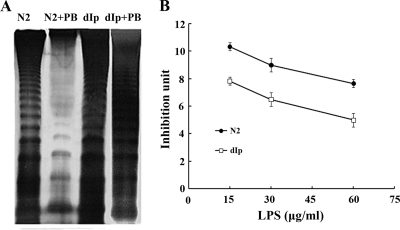
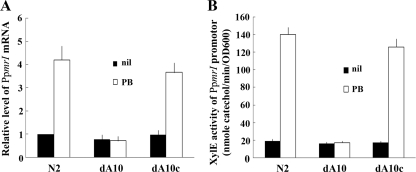
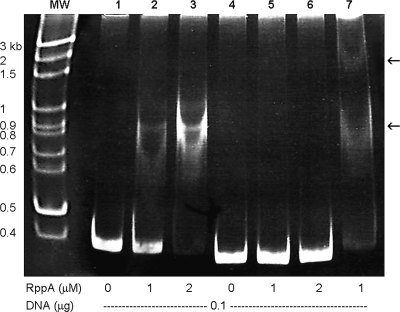
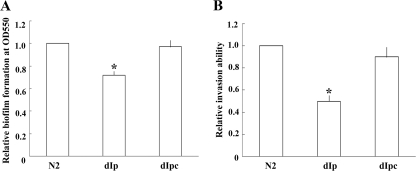
Proteus mirabilis is naturally resistant to polymyxin B (PB). To investigate the underlying mechanisms, Tn5 mutagenesis was performed, and a mutant exhibiting increased PB susceptibility was isolated. The mutant was found to have Tn5 inserted into the PpmrI (Proteus pmrI) gene, a gene which may encode a UDP-glucuronic acid decarboxylase. In other bacteria, pmrI belongs to the seven-gene pmrF operon, which is involved in lipopolysaccharide (LPS) modification. While the PpmrI knockout mutant had a wild-type LPS profile and produced amounts of LPS similar to those produced by the wild type, LPS of the knockout mutant had higher PB-binding activity than that of the wild type. PB could induce alterations of LPS in the wild type but not in the PpmrI knockout mutant. Moreover, the PpmrI knockout mutant exhibited decreased abilities in biofilm formation and urothelial cell invasion. Complementation of the PpmrI mutant with the full-length PpmrI gene led to restoration of the wild-type phenotypic traits. Previously we identified RppA, a response regulator of the bacterial two-component system, as a regulator of PB susceptibility and virulence factor expression in P. mirabilis. Here we showed that RppA could mediate the induction of PpmrI expression by PB. An electrophoretic mobility shift assay further demonstrated that RppA could bind directly to the putative PpmrI promoter. Together, these results provide a new insight into the regulatory mechanism underlying PB resistance and virulence expression in P. mirabilis.
Figures
Similar articles
-
Role of RppA in the regulation of polymyxin b susceptibility, swarming, and virulence factor expression in Proteus mirabilis.Infect Immun. 2008 May;76(5):2051-62. doi: 10.1128/IAI.01557-07. Epub 2008 Mar 3. Infect Immun. 2008. PMID: 18316383 Free PMC article.
-
Characterization of UDP-glucose dehydrogenase and UDP-glucose pyrophosphorylase mutants of Proteus mirabilis: defectiveness in polymyxin B resistance, swarming, and virulence.Antimicrob Agents Chemother. 2010 May;54(5):2000-9. doi: 10.1128/AAC.01384-09. Epub 2010 Feb 16. Antimicrob Agents Chemother. 2010. PMID: 20160049 Free PMC article.
-
10'(Z),13'(E)-heptadecadienylhydroquinone inhibits swarming and virulence factors and increases polymyxin B susceptibility in Proteus mirabilis.PLoS One. 2012;7(9):e45563. doi: 10.1371/journal.pone.0045563. Epub 2012 Sep 20. PLoS One. 2012. PMID: 23029100 Free PMC article.
-
The chemical structure of the lipopolysaccharide of A Rc-type mutant of Proteus mirabilis lacking 4-amino-4-deoxy-L arabinose and its susceptibility towards polymyxin B.Adv Exp Med Biol. 1990;256:121-6. doi: 10.1007/978-1-4757-5140-6_7. Adv Exp Med Biol. 1990. PMID: 2158212 Review. No abstract available.
-
Swarmer cell differentiation in Proteus mirabilis.Environ Microbiol. 2005 Aug;7(8):1065-73. doi: 10.1111/j.1462-2920.2005.00806.x. Environ Microbiol. 2005. PMID: 16011745 Review.
Cited by
-
A cyclic adenosine monophosphate response element-binding protein inhibitor enhances the antibacterial activity of polymyxin B by inhibiting the ATP hydrolyzation activity of CrrB.Front Pharmacol. 2022 Sep 6;13:949869. doi: 10.3389/fphar.2022.949869. eCollection 2022. Front Pharmacol. 2022. PMID: 36147339 Free PMC article.
-
The RNA chaperone Hfq is involved in stress tolerance and virulence in uropathogenic Proteus mirabilis.PLoS One. 2014 Jan 15;9(1):e85626. doi: 10.1371/journal.pone.0085626. eCollection 2014. PLoS One. 2014. PMID: 24454905 Free PMC article.
-
Identification and Functional Analysis of ncRNAs Regulating Intrinsic Polymyxin Resistance in Foodborne Proteus vulgaris.Microorganisms. 2024 Aug 13;12(8):1661. doi: 10.3390/microorganisms12081661. Microorganisms. 2024. PMID: 39203505 Free PMC article.
-
Polymyxin: Alternative Mechanisms of Action and Resistance.Cold Spring Harb Perspect Med. 2016 Oct 3;6(10):a025288. doi: 10.1101/cshperspect.a025288. Cold Spring Harb Perspect Med. 2016. PMID: 27503996 Free PMC article. Review.
-
Isolation, biological and whole genome characteristics of a Proteus mirabilis bacteriophage strain.BMC Microbiol. 2023 Aug 8;23(1):215. doi: 10.1186/s12866-023-02960-4. BMC Microbiol. 2023. PMID: 37553593 Free PMC article.
References
-
- Bader, M. W., W. W. Navarre, W. Shiau, H. Nikaido, J. G. Frye, M. McClelland, F. C. Fang, and S. I. Miller. 2003. Regulation of Salmonella typhimurium virulence gene expression by cationic antimicrobial peptides. Mol. Microbiol. 50:219-230. - PubMed
-
- Bader, M. W., S. Sanowar, M. E. Daley, A. R. Schneider, U. Cho, W. Xu, R. E. Klevit, H. Le Moual, and S. I. Miller. 2005. Recognition of antimicrobial peptides by a bacterial sensor kinase. Cell 122:461-472. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
