

Intrahepatic IL-10 maintains NKG2A+Ly49- liver NK cells in a functionally hyporesponsive state
- PMID: 20124099
- PMCID: PMC2885840
- DOI: 10.4049/jimmunol.0901362
Intrahepatic IL-10 maintains NKG2A+Ly49- liver NK cells in a functionally hyporesponsive state
Abstract
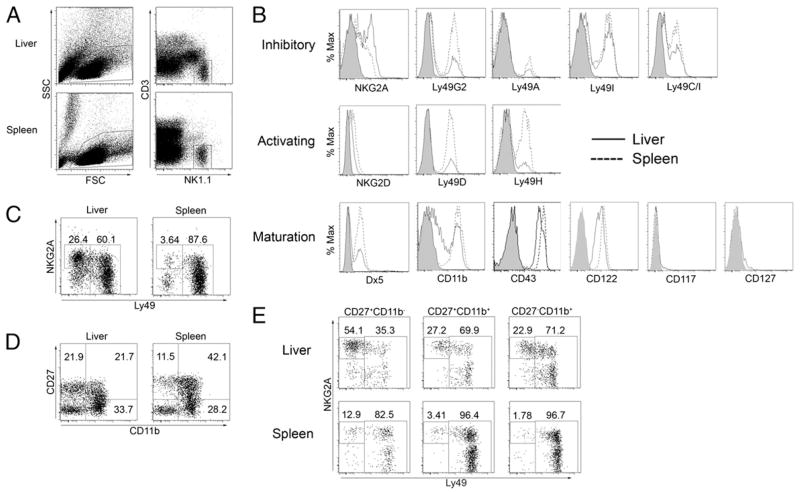
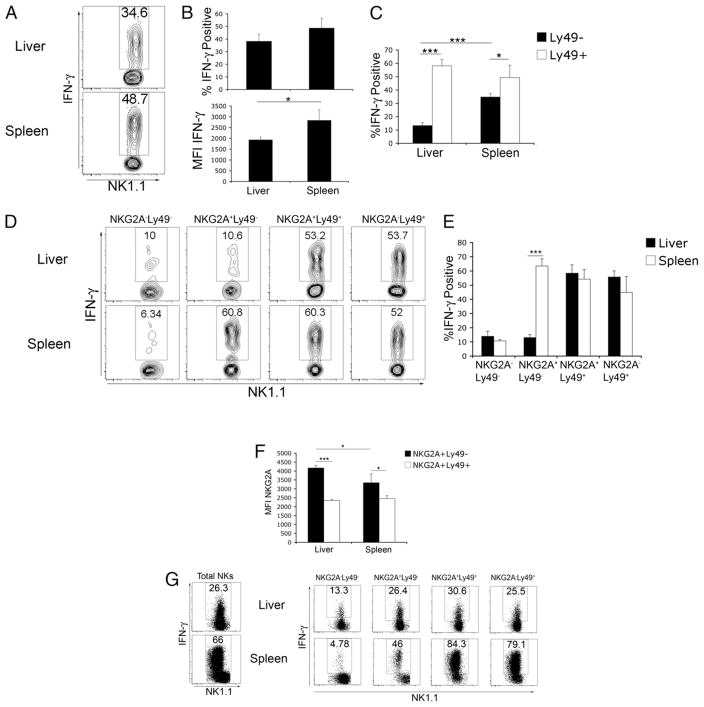
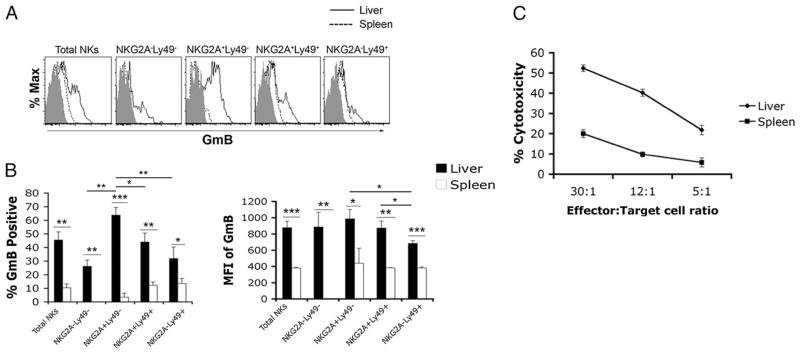
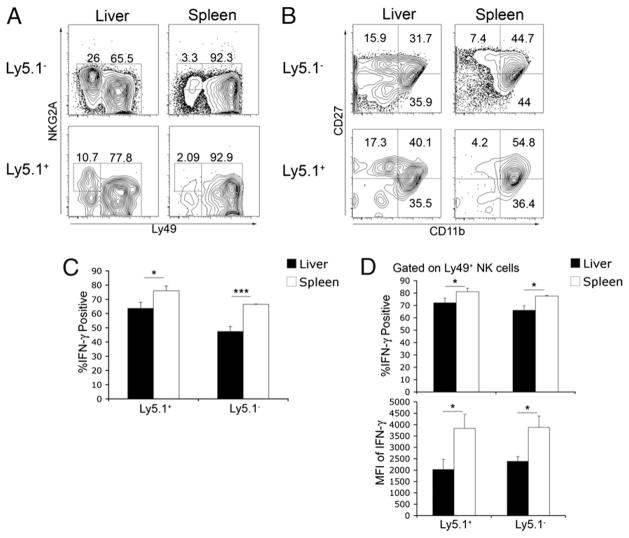
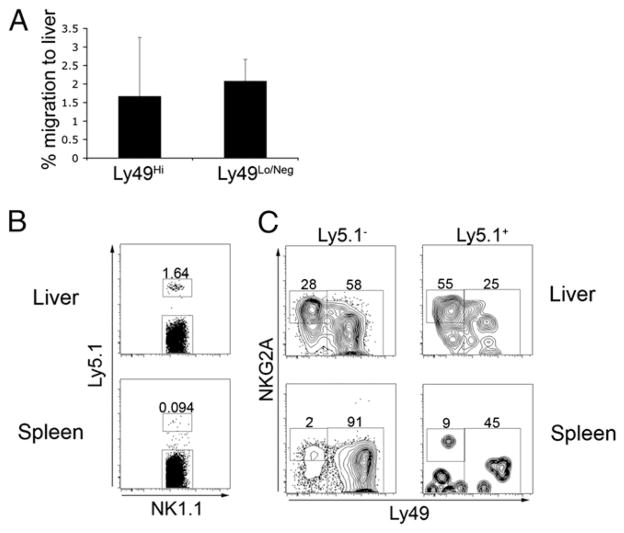
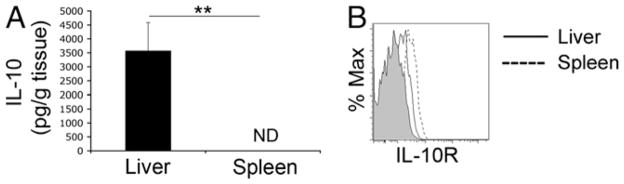
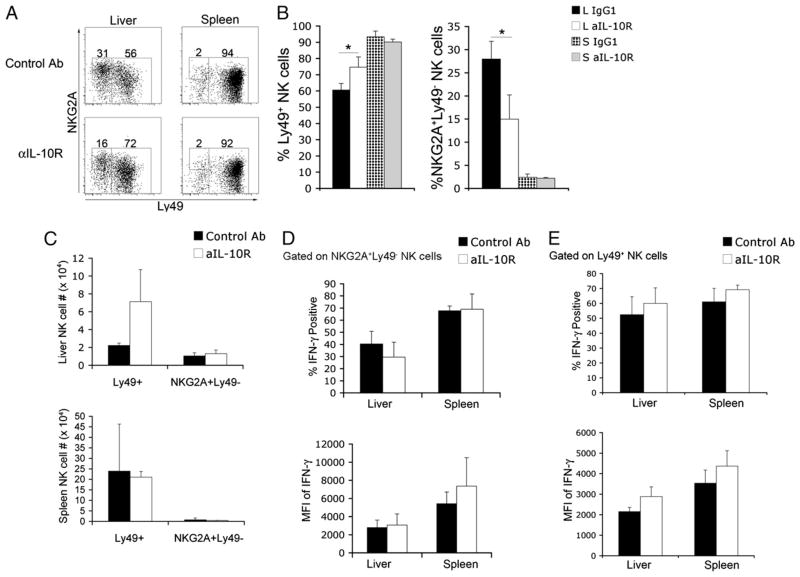
The tolerogenic nature of the liver allows daily exposure to gut-derived foreign Ags without causing inflammation, but it may facilitate persistent infection in the liver. NK cells play a central role in innate immunity, as well as in shaping the adaptive immune response. We hypothesized that the naive mouse liver maintains intrahepatic NK cells in a functionally hyporesponsive state. Compared with splenic NK cells, liver NK cells displayed a dampened IFN-gamma response to IL-12/IL-18 stimulation. Importantly, the liver contains a significant population of functionally hyporesponsive NK cells that express high levels of the inhibitory receptor NKG2A and lack expression of MHC class I-binding Ly49 receptors. Adoptively transferred splenic NK cells that migrate to the liver displayed phenotypic and functional changes, suggesting that the liver environment modifies NK cell receptor expression and functional responsiveness. Notably, IL-10 is present at high levels within the liver, and in vivo blockade of IL-10R resulted in a decreased percentage of intrahepatic NKG2A(+)Ly49(-) NK cells. These data suggest that the liver environment regulates NK cell receptor expression and that IL-10 contributes to the regulation of liver NK cells, in part, by maintaining a greater percentage of the hyporesponsive NKG2A(+)Ly49(-) NK cells in the liver.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures
References
-
- Andrews DM, Scalzo AA, Yokoyama WM, Smyth MJ, Degli-Esposti MA. Functional interactions between dendritic cells and NK cells during viral infection. Nat Immunol. 2003;4:175–181. - PubMed
-
- Martín-Fontecha A, Thomsen LL, Brett S, Gerard C, Lipp M, Lanzavecchia A, Sallusto F. Induced recruitment of NK cells to lymph nodes provides IFN-gamma for T(H)1 priming. Nat Immunol. 2004;5:1260–1265. - PubMed
-
- Kim S, Iizuka K, Kang HS, Dokun A, French AR, Greco S, Yokoyama WM. In vivo developmental stages in murine natural killer cell maturation. Nat Immunol. 2002;3:523–528. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
