

Differential expression of claudin tight junction proteins in the human cortical nephron
- PMID: 20124215
- PMCID: PMC2891746
- DOI: 10.1093/ndt/gfq006
Differential expression of claudin tight junction proteins in the human cortical nephron
Abstract
Background: In renal tubules, paracellular permeability is tightly controlled to facilitate solute absorption and urinary concentration and is regulated by tight junctions, which incorporate claudin proteins. There is very limited information confirming the localization of these proteins in the human renal cortex. Most data is inferred from mouse, bovine and rabbit studies and differences exist between mouse and other species.
Methods: A survey of claudin staining was performed on human kidney cortex embedded in glycolmethacrylate resin to enhance tissue morphology and facilitate the cutting of 2 microm serial sections.
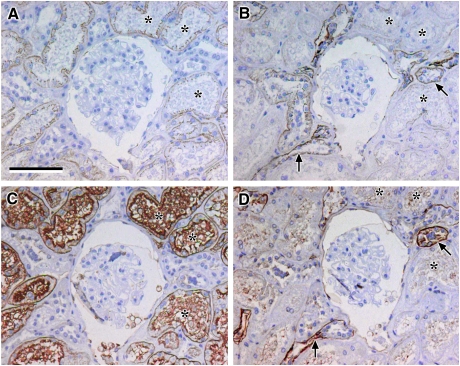
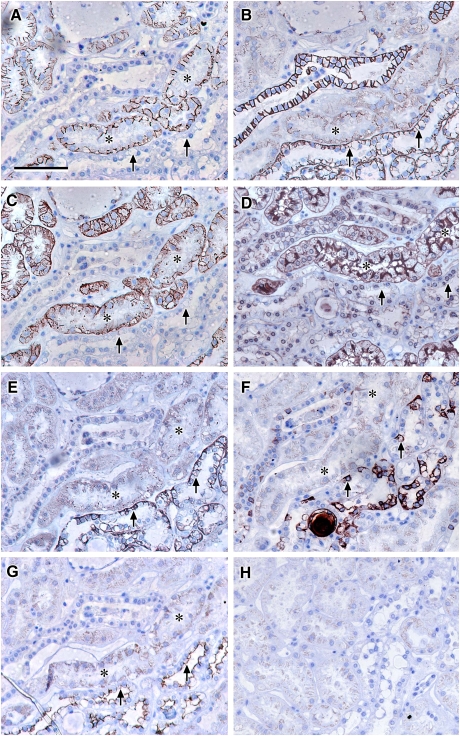
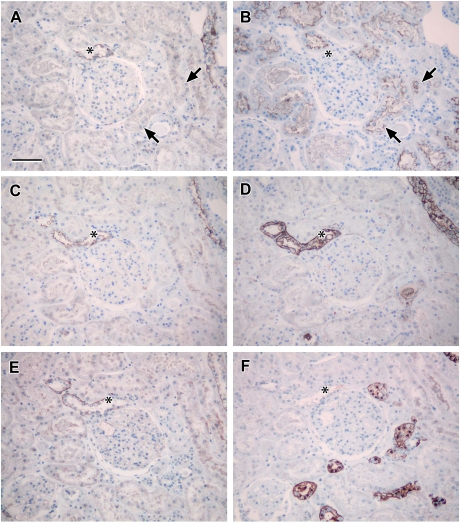
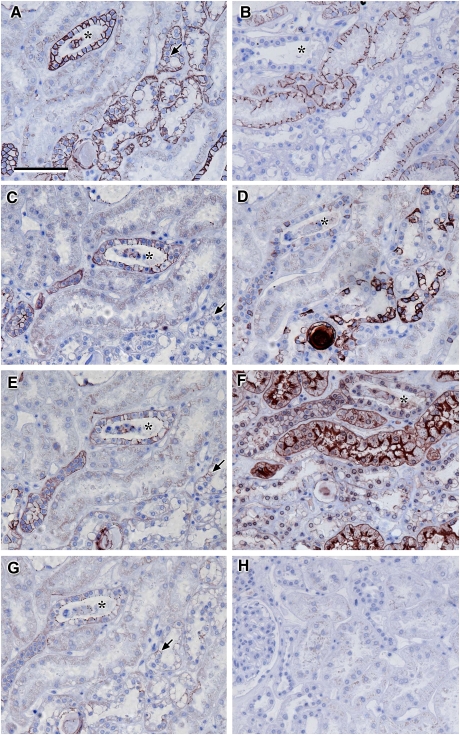
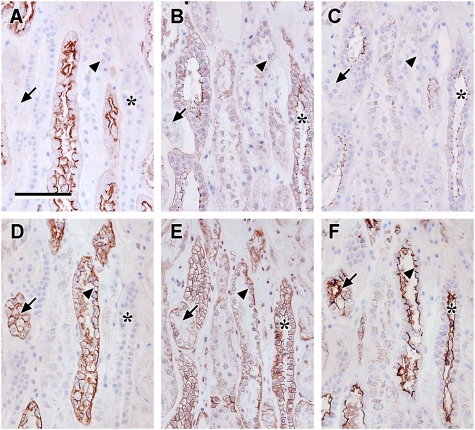
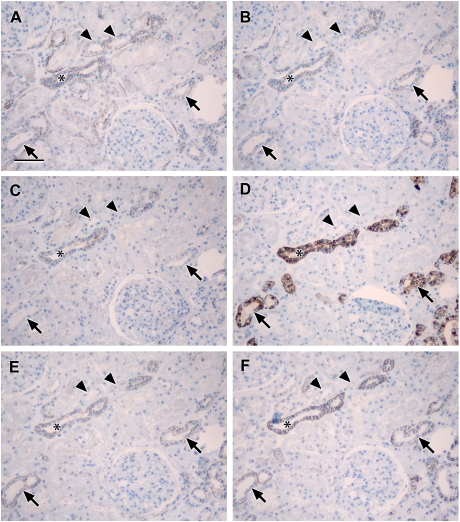
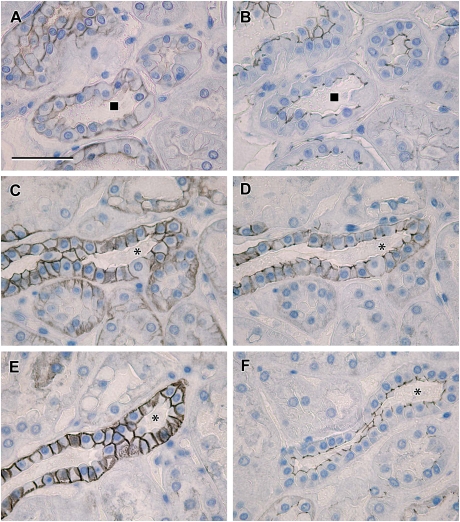
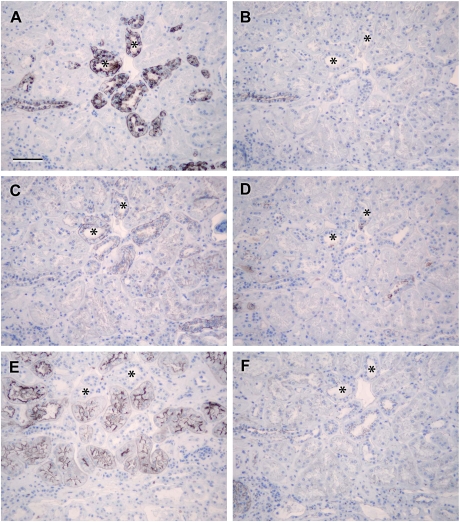
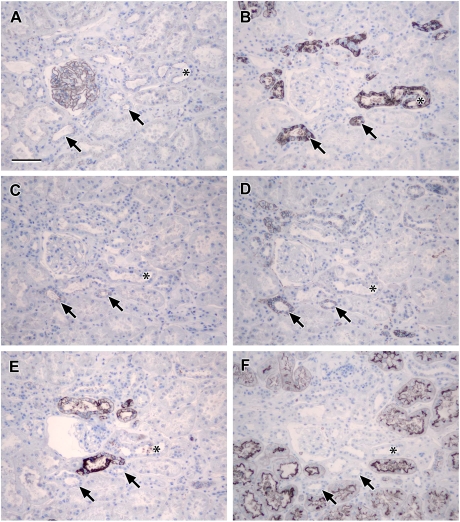
Results: Claudin-2, -10 and -11 antibodies labelled renal tubular epithelial cells, correlating with Lotus tetragonolobus and N-cadherin positive proximal tubules. Claudin-3, -10, -11 and -16 antibodies strongly stained a population of tubules that were positive for Tamm Horsfall protein on adjacent sections, confirming expression in the thick ascending limb of the Loop of Henle. Claudin-3, -4 and -8 antibodies reacted with tubules that correlated with the distal nephron markers, E-cadherin, epithelial membrane antigen and Dolichos biflorus and claudin-3, -4, -7 and -8 with the distal tubule marker, calbindin, and the collecting duct marker, aquaporin-2. Claudin-14 was localized in distal convoluted tubules, correlating positively with calbindin but negatively with aquaporin-2, whereas claudin-1 staining was identified in the parietal epithelium of Bowman's capsule, distal convoluted tubule and collecting duct. Cellular and tight junction localization of claudin staining in renal tubules was heterogeneous and is discussed.
Conclusions: Complex variation in the expression of human claudins likely determines paracellular permeability in the kidney. Altered claudin expression may influence pathologies involving abnormalities of absorption.
Figures
References
-
- Rector FC., Jr Sodium, bicarbonate, and chloride absorption by the proximal tubule. Am J Physiol. 1983;244:F461–F471. - PubMed
-
- Berry CA, Rector FC., Jr Mechanism of proximal NaCl reabsorption in the proximal tubule of the mammalian kidney. Semin Nephrol. 1991;11:86–97. - PubMed
-
- Aronson PS, Giebisch G. Mechanisms of chloride transport in the proximal tubule. Am J Physiol. 1997;273:F179–F192. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
