

Identification and validation of novel spinophilin-associated proteins in rodent striatum using an enhanced ex vivo shotgun proteomics approach
- PMID: 20124353
- PMCID: PMC2877984
- DOI: 10.1074/mcp.M900387-MCP200
Identification and validation of novel spinophilin-associated proteins in rodent striatum using an enhanced ex vivo shotgun proteomics approach
Abstract
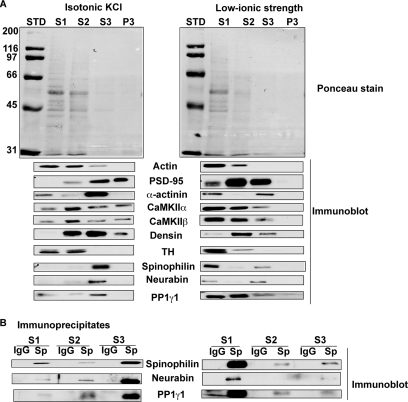
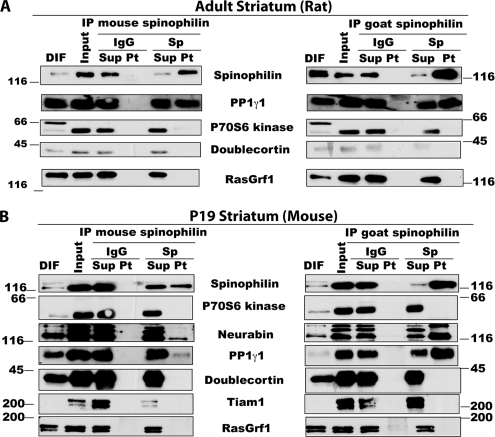
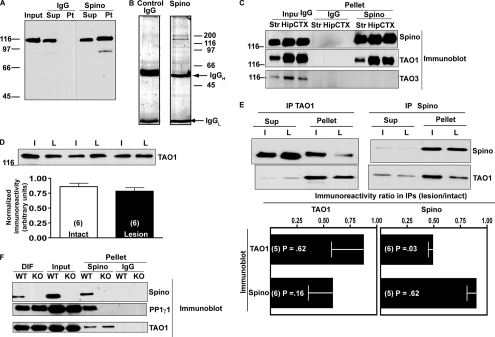
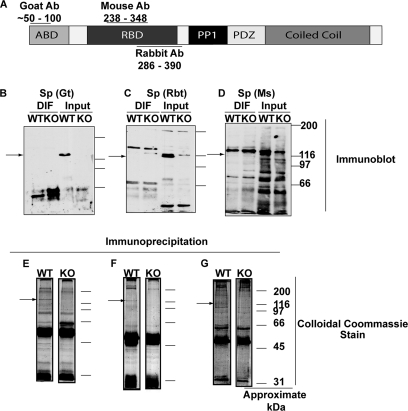
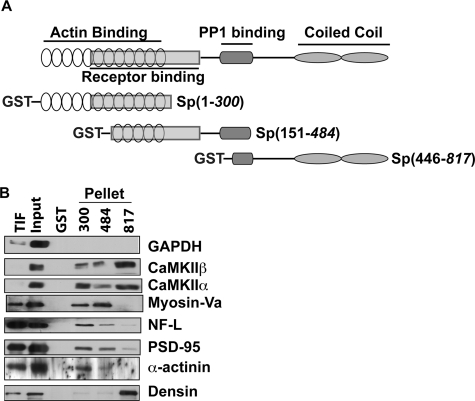
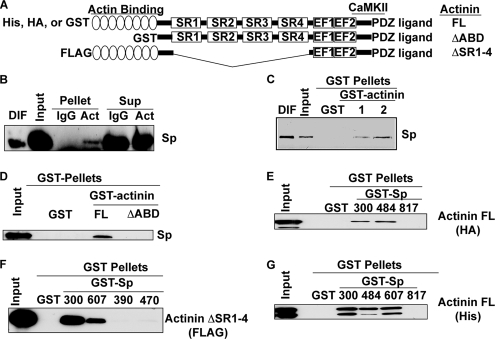
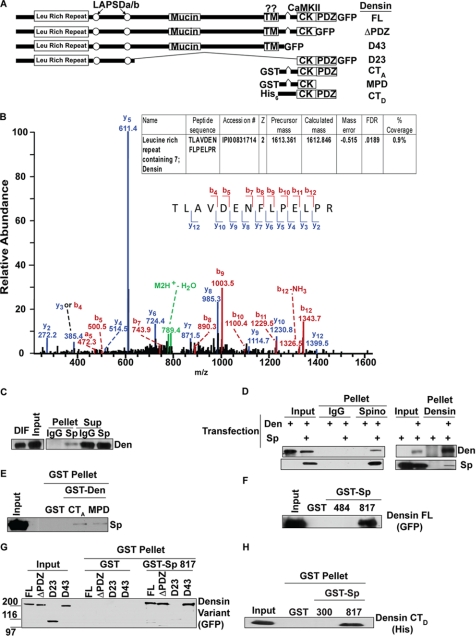
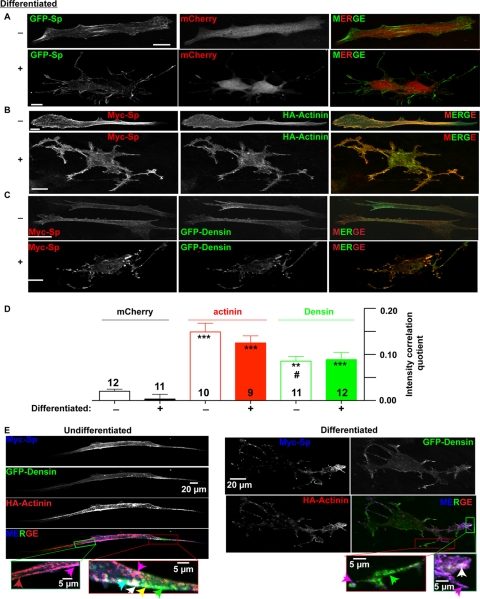
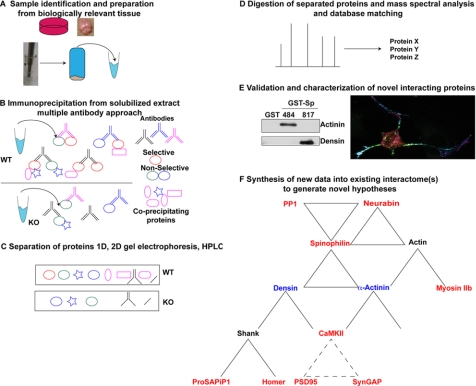
Spinophilin regulates excitatory postsynaptic function and morphology during development by virtue of its interactions with filamentous actin, protein phosphatase 1, and a plethora of additional signaling proteins. To provide insight into the roles of spinophilin in mature brain, we characterized the spinophilin interactome in subcellular fractions solubilized from adult rodent striatum by using a shotgun proteomics approach to identify proteins in spinophilin immune complexes. Initial analyses of samples generated using a mouse spinophilin antibody detected 23 proteins that were not present in an IgG control sample; however, 12 of these proteins were detected in complexes isolated from spinophilin knock-out tissue. A second screen using two different spinophilin antibodies and either knock-out or IgG controls identified a total of 125 proteins. The probability of each protein being specifically associated with spinophilin in each sample was calculated, and proteins were ranked according to a chi(2) analysis of the probabilities from analyses of multiple samples. Spinophilin and the known associated proteins neurabin and multiple isoforms of protein phosphatase 1 were specifically detected. Multiple, novel, spinophilin-associated proteins (myosin Va, calcium/calmodulin-dependent protein kinase II, neurofilament light polypeptide, postsynaptic density 95, alpha-actinin, and densin) were then shown to interact with GST fusion proteins containing fragments of spinophilin. Additional biochemical and transfected cell imaging studies showed that alpha-actinin and densin directly interact with residues 151-300 and 446-817, respectively, of spinophilin. Taken together, we have developed a multi-antibody, shotgun proteomics approach to characterize protein interactomes in native tissues, delineating the importance of knock-out tissue controls and providing novel insights into the nature and function of the spinophilin interactome in mature striatum.
Figures
References
-
- Kim E., Sheng M. (2004) PDZ domain proteins of synapses. Nat. Rev. Neurosci 5, 771–781 - PubMed
-
- Kennedy M. B., Beale H. C., Carlisle H. J., Washburn L. R. (2005) Integration of biochemical signalling in spines. Nat. Rev. Neurosci 6, 423–434 - PubMed
-
- Dindot S. V., Antalffy B. A., Bhattacharjee M. B., Beaudet A. L. (2008) The Angelman syndrome ubiquitin ligase localizes to the synapse and nucleus, and maternal deficiency results in abnormal dendritic spine morphology. Hum. Mol. Genet 17, 111–118 - PubMed
-
- Ingham C. A., Hood S. H., Arbuthnott G. W. (1989) Spine density on neostriatal neurones changes with 6-hydroxydopamine lesions and with age. Brain Res 503, 334–338 - PubMed
-
- Ingham C. A., Hood S. H., van Maldegem B., Weenink A., Arbuthnott G. W. (1993) Morphological changes in the rat neostriatum after unilateral 6-hydroxydopamine injections into the nigrostriatal pathway. Exp. Brain Res 93, 17–27 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01-MH63232/MH/NIMH NIH HHS/United States
- P01 NS044282/NS/NINDS NIH HHS/United States
- R01 CA126218/CA/NCI NIH HHS/United States
- DK58404/DK/NIDDK NIH HHS/United States
- P30 HD015052/HD/NICHD NIH HHS/United States
- EY08126/EY/NEI NIH HHS/United States
- P30 EY008126/EY/NEI NIH HHS/United States
- P01-NS044282/NS/NINDS NIH HHS/United States
- R01 MH063232/MH/NIMH NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- DK20593/DK/NIDDK NIH HHS/United States
- F31 NS061537/NS/NINDS NIH HHS/United States
- U24 DK059637/DK/NIDDK NIH HHS/United States
- P60 DK020593/DK/NIDDK NIH HHS/United States
- P30CA68485/CA/NCI NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- T32 MH065215/MH/NIMH NIH HHS/United States
- T32-MH65215/MH/NIMH NIH HHS/United States
- DK59637/DK/NIDDK NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- NS061537/NS/NINDS NIH HHS/United States
- CA68485/CA/NCI NIH HHS/United States
- HD15052/HD/NICHD NIH HHS/United States
- 1R01CA126218/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
