

High-mobility group box 1 is involved in the initial events of early loss of transplanted islets in mice
- PMID: 20124731
- PMCID: PMC2827961
- DOI: 10.1172/JCI41360
High-mobility group box 1 is involved in the initial events of early loss of transplanted islets in mice
Abstract
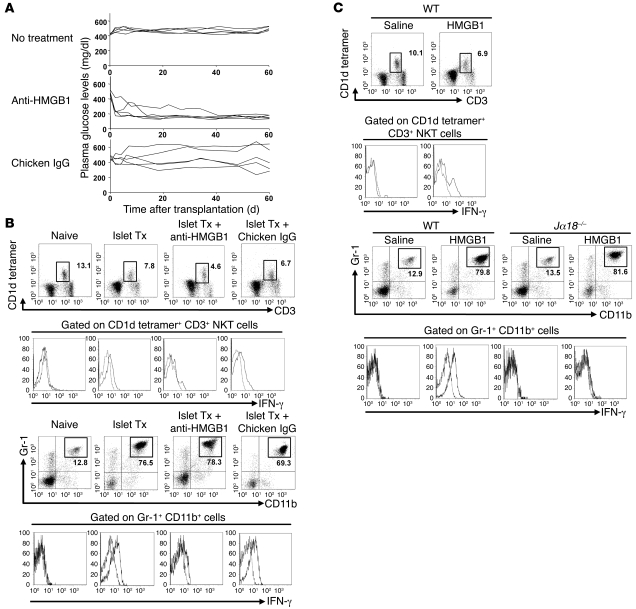
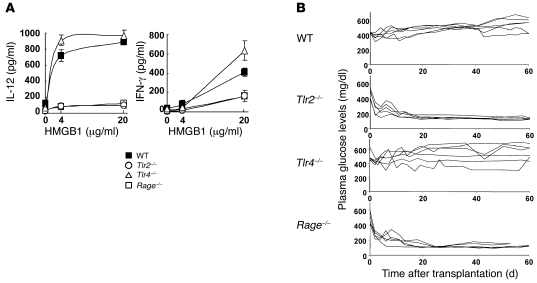
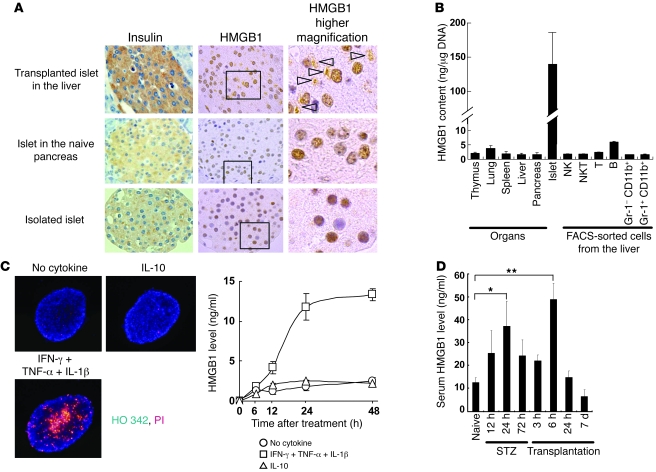
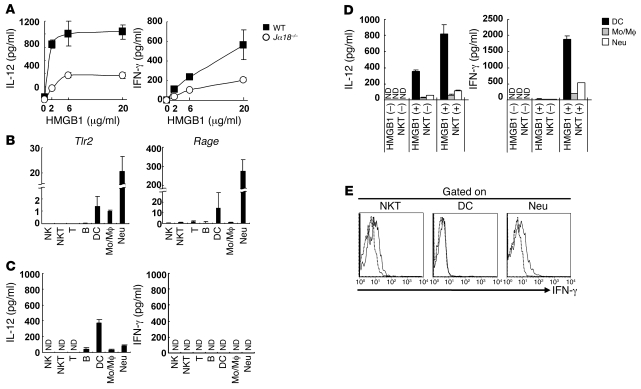
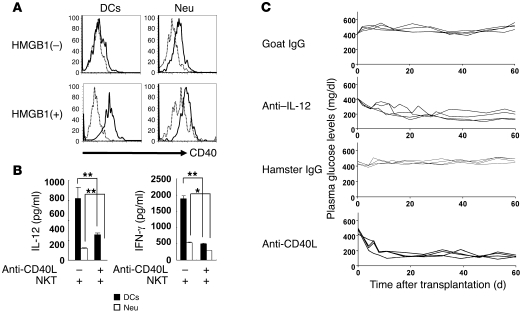
Islet transplantation for the treatment of type 1 diabetes mellitus is limited in its clinical application mainly due to early loss of the transplanted islets, resulting in low transplantation efficiency. NKT cell-dependent IFN-gamma production by Gr-1(+)CD11b(+) cells is essential for this loss, but the upstream events in the process remain undetermined. Here, we have demonstrated that high-mobility group box 1 (HMGB1) plays a crucial role in the initial events of early loss of transplanted islets in a mouse model of diabetes. Pancreatic islets contained abundant HMGB1, which was released into the circulation soon after islet transplantation into the liver. Treatment with an HMGB1-specific antibody prevented the early islet graft loss and inhibited IFN-gamma production by NKT cells and Gr-1(+)CD11b(+) cells. Moreover, mice lacking either of the known HMGB1 receptors TLR2 or receptor for advanced glycation end products (RAGE), but not the known HMGB1 receptor TLR4, failed to exhibit early islet graft loss. Mechanistically, HMGB1 stimulated hepatic mononuclear cells (MNCs) in vivo and in vitro; in particular, it upregulated CD40 expression and enhanced IL-12 production by DCs, leading to NKT cell activation and subsequent NKT cell-dependent augmented IFN-gamma production by Gr-1(+)CD11b(+) cells. Thus, treatment with either IL-12- or CD40L-specific antibody prevented the early islet graft loss. These findings indicate that the HMGB1-mediated pathway eliciting early islet loss is a potential target for intervention to improve the efficiency of islet transplantation.
Figures
Similar articles
-
HMGB1-Mediated Early Loss of Transplanted Islets Is Prevented by Anti-IL-6R Antibody in Mice.Pancreas. 2015 Jan;44(1):166-71. doi: 10.1097/MPA.0000000000000188. Pancreas. 2015. PMID: 25058889
-
Islet-expressed TLR2 and TLR4 sense injury and mediate early graft failure after transplantation.Eur J Immunol. 2010 Oct;40(10):2914-24. doi: 10.1002/eji.201040601. Eur J Immunol. 2010. PMID: 20809521 Free PMC article.
-
Prevention of high-mobility group box 1-mediated early loss of transplanted mouse islets in the liver by antithrombin III.Transplantation. 2012 May 27;93(10):983-8. doi: 10.1097/TP.0b013e31824d3508. Transplantation. 2012. PMID: 22446936
-
HMGB1, an innate alarmin, in the pathogenesis of type 1 diabetes.Int J Clin Exp Pathol. 2009 Sep 8;3(1):24-38. Int J Clin Exp Pathol. 2009. PMID: 19918326 Free PMC article. Review.
-
Cell Surface Engineering for Regulation of Immune Reactions in Cell Therapy.Adv Exp Med Biol. 2015;865:189-209. doi: 10.1007/978-3-319-18603-0_12. Adv Exp Med Biol. 2015. PMID: 26306451 Review.
Cited by
-
Withaferin A inhibits pro-inflammatory cytokine-induced damage to islets in culture and following transplantation.Diabetologia. 2013 Apr;56(4):814-24. doi: 10.1007/s00125-012-2813-9. Epub 2013 Jan 15. Diabetologia. 2013. PMID: 23318585
-
Evolution of β-Cell Replacement Therapy in Diabetes Mellitus: Islet Cell Transplantation.J Transplant. 2011;2011:247959. doi: 10.1155/2011/247959. Epub 2011 Oct 15. J Transplant. 2011. PMID: 22013505 Free PMC article.
-
Relationship between HMGB1 and PAI-1 after allogeneic hematopoietic stem cell transplantation.J Blood Med. 2016 Jan 18;7:1-4. doi: 10.2147/JBM.S93008. eCollection 2016. J Blood Med. 2016. PMID: 26848281 Free PMC article.
-
The Spleen Is an Ideal Site for Inducing Transplanted Islet Graft Expansion in Mice.PLoS One. 2017 Jan 30;12(1):e0170899. doi: 10.1371/journal.pone.0170899. eCollection 2017. PLoS One. 2017. PMID: 28135283 Free PMC article.
-
Ex vivo Pretreatment of Islets with Mitomycin C: Reduction in Immunogenic Potential of Islets by Suppressing Secretion of Multiple Chemotactic Factors.Cell Transplant. 2017 Aug;26(8):1392-1404. doi: 10.1177/0963689717721233. Cell Transplant. 2017. PMID: 28901184 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
