

Role of plastid protein phosphatase TAP38 in LHCII dephosphorylation and thylakoid electron flow
- PMID: 20126264
- PMCID: PMC2811158
- DOI: 10.1371/journal.pbio.1000288
Role of plastid protein phosphatase TAP38 in LHCII dephosphorylation and thylakoid electron flow
Abstract
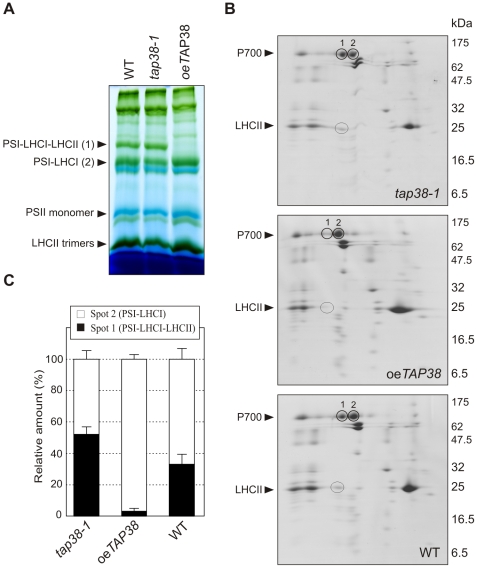
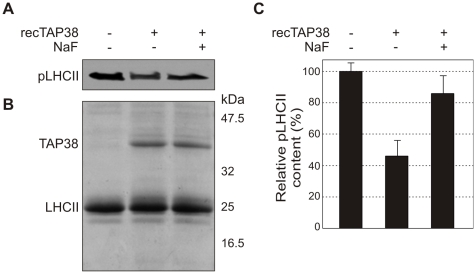
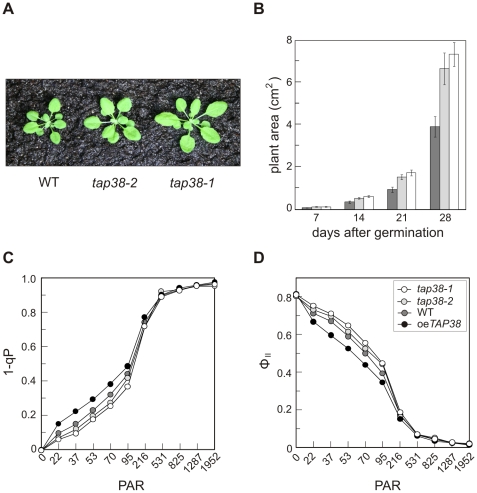
Short-term changes in illumination elicit alterations in thylakoid protein phosphorylation and reorganization of the photosynthetic machinery. Phosphorylation of LHCII, the light-harvesting complex of photosystem II, facilitates its relocation to photosystem I and permits excitation energy redistribution between the photosystems (state transitions). The protein kinase STN7 is required for LHCII phosphorylation and state transitions in the flowering plant Arabidopsis thaliana. LHCII phosphorylation is reversible, but extensive efforts to identify the protein phosphatase(s) that dephosphorylate LHCII have been unsuccessful. Here, we show that the thylakoid-associated phosphatase TAP38 is required for LHCII dephosphorylation and for the transition from state 2 to state 1 in A. thaliana. In tap38 mutants, thylakoid electron flow is enhanced, resulting in more rapid growth under constant low-light regimes. TAP38 gene overexpression markedly decreases LHCII phosphorylation and inhibits state 1-->2 transition, thus mimicking the stn7 phenotype. Furthermore, the recombinant TAP38 protein is able, in an in vitro assay, to directly dephosphorylate LHCII. The dependence of LHCII dephosphorylation upon TAP38 dosage, together with the in vitro TAP38-mediated dephosphorylation of LHCII, suggests that TAP38 directly acts on LHCII. Although reversible phosphorylation of LHCII and state transitions are crucial for plant fitness under natural light conditions, LHCII hyperphosphorylation associated with an arrest of photosynthesis in state 2 due to inactivation of TAP38 improves photosynthetic performance and plant growth under state 2-favoring light conditions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Plants Actively Avoid State Transitions upon Changes in Light Intensity: Role of Light-Harvesting Complex II Protein Dephosphorylation in High Light.Plant Physiol. 2015 Jun;168(2):721-34. doi: 10.1104/pp.15.00488. Epub 2015 Apr 22. Plant Physiol. 2015. PMID: 25902812 Free PMC article.
-
Dynamics of reversible protein phosphorylation in thylakoids of flowering plants: the roles of STN7, STN8 and TAP38.Biochim Biophys Acta. 2011 Aug;1807(8):887-96. doi: 10.1016/j.bbabio.2010.08.002. Epub 2010 Aug 20. Biochim Biophys Acta. 2011. PMID: 20728426 Review.
-
Structural Mechanism Underlying the Specific Recognition between the Arabidopsis State-Transition Phosphatase TAP38/PPH1 and Phosphorylated Light-Harvesting Complex Protein Lhcb1.Plant Cell. 2015 Apr;27(4):1113-27. doi: 10.1105/tpc.15.00102. Epub 2015 Apr 17. Plant Cell. 2015. PMID: 25888588 Free PMC article.
-
STN7 is not essential for developmental acclimation of Arabidopsis to light intensity.Plant J. 2023 Jun;114(6):1458-1474. doi: 10.1111/tpj.16204. Epub 2023 Apr 11. Plant J. 2023. PMID: 36960687 Free PMC article.
-
Thylakoid Protein Phosphorylation in Chloroplasts.Plant Cell Physiol. 2021 Oct 29;62(7):1094-1107. doi: 10.1093/pcp/pcab043. Plant Cell Physiol. 2021. PMID: 33768241 Review.
Cited by
-
Phylogenetic viewpoints on regulation of light harvesting and electron transport in eukaryotic photosynthetic organisms.Planta. 2013 Feb;237(2):399-412. doi: 10.1007/s00425-012-1744-5. Epub 2012 Sep 13. Planta. 2013. PMID: 22971817 Review.
-
Combinatory actions of CP29 phosphorylation by STN7 and stability regulate leaf age-dependent disassembly of photosynthetic complexes.Sci Rep. 2020 Jun 24;10(1):10267. doi: 10.1038/s41598-020-67213-0. Sci Rep. 2020. PMID: 32581255 Free PMC article.
-
Single chloroplast in folio imaging sheds light on photosystem energy redistribution during state transitions.Plant Physiol. 2023 Feb 12;191(2):1186-1198. doi: 10.1093/plphys/kiac561. Plant Physiol. 2023. PMID: 36478277 Free PMC article.
-
State Transition Regulation in Chlamydomonas reinhardtii.Plant Physiol. 2020 Aug;183(4):1418-1419. doi: 10.1104/pp.20.00814. Plant Physiol. 2020. PMID: 32747490 Free PMC article. No abstract available.
-
The major thylakoid protein kinases STN7 and STN8 revisited: effects of altered STN8 levels and regulatory specificities of the STN kinases.Front Plant Sci. 2013 Oct 21;4:417. doi: 10.3389/fpls.2013.00417. eCollection 2013. Front Plant Sci. 2013. PMID: 24151498 Free PMC article.
References
-
- Allen J. F, Forsberg J. Molecular recognition in thylakoid structure and function. Trends Plant Sci. 2001;6:317–326. - PubMed
-
- Eberhard S, Finazzi G, Wollman F. A. The dynamics of photosynthesis. Annu Rev Genet. 2008;42:463–515. - PubMed
-
- Haldrup A, Jensen P. E, Lunde C, Scheller H. V. Balance of power: a view of the mechanism of photosynthetic state transitions. Trends Plant Sci. 2001;6:301–305. - PubMed
-
- Rochaix J. D. Role of thylakoid protein kinases in photosynthetic acclimation. FEBS Lett. 2007;581:2768–2775. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
