

Agroinfiltration reduces ABA levels and suppresses Pseudomonas syringae-elicited salicylic acid production in Nicotiana tabacum
- PMID: 20126459
- PMCID: PMC2813289
- DOI: 10.1371/journal.pone.0008977
Agroinfiltration reduces ABA levels and suppresses Pseudomonas syringae-elicited salicylic acid production in Nicotiana tabacum
Abstract
Background: Agrobacterium tumefaciens strain GV3101 (pMP90) is widely used in transient gene expression assays, including assays to study pathogen effectors and plant disease resistance mechanisms. However, inoculation of A. tumefaciens GV3101 into Nicotiana tabacum (tobacco) leaves prior to infiltration with pathogenic and non-host strains of Pseudomonas syringae results in suppression of macroscopic symptoms when compared with leaves pre-treated with a buffer control.
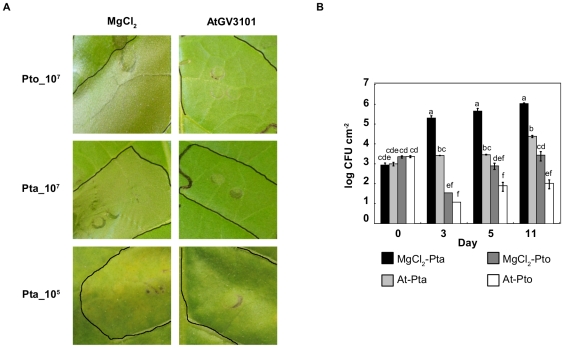
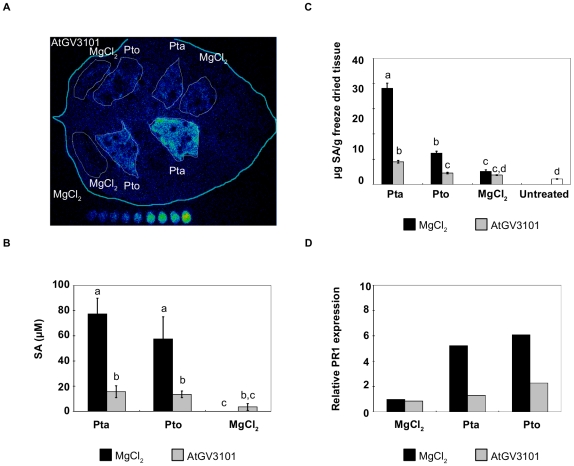
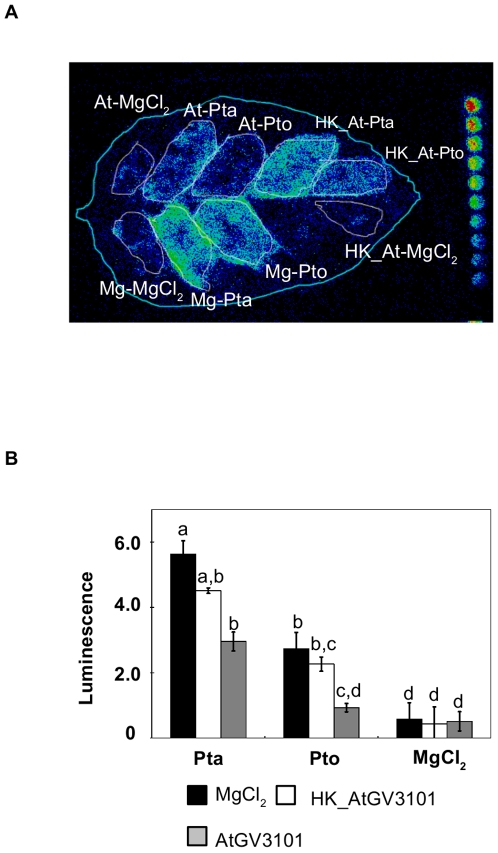
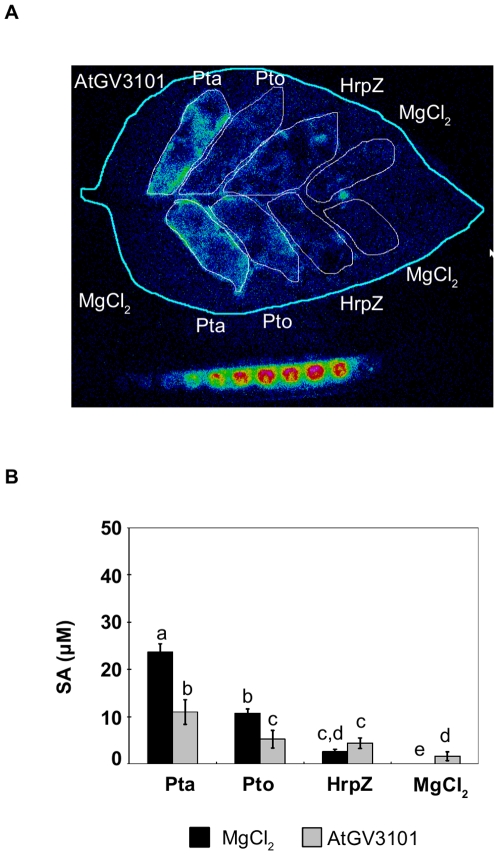
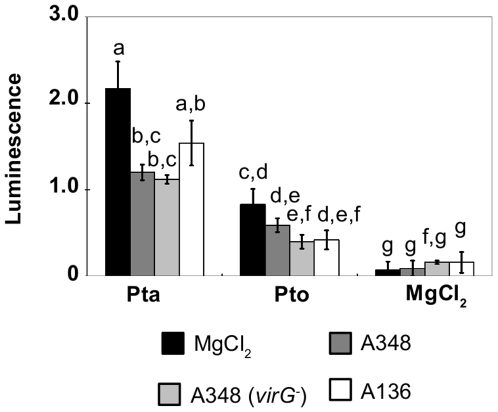
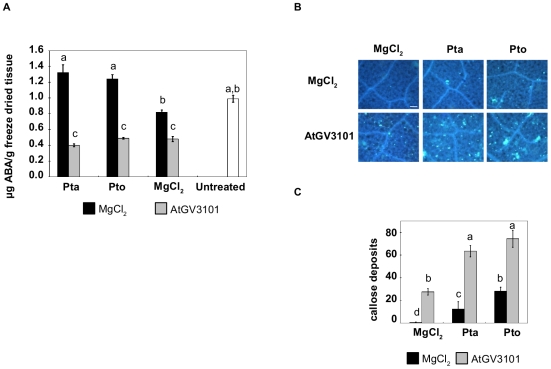
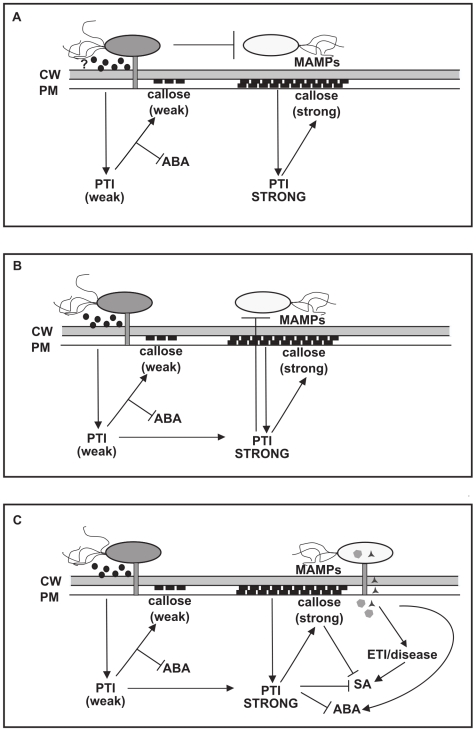
Methodology/findings: To gain further insight into the mechanistic basis of symptom suppression by A. tumefaciens we examined the effect of pre-treatment with A. tumefaciens on the growth of P. syringae, the production of the plant signalling molecules salicylic acid (SA) and abscisic acid (ABA), and the presence of callose deposits. Pre-treatment with A. tumefaciens reduced ABA levels, P. syringae multiplication and P. syringae-elicited SA and ABA production, but promoted increased callose deposition. However, pre-treatment with A. tumefaciens did not suppress necrosis or SA production in leaves inoculated with the elicitor HrpZ.
Conclusions/significance: Collectively, these results show that inoculation of N. tabacum leaves with A. tumefaciens alters plant hormone levels and plant defence responses to P. syringae, and demonstrate that researchers should consider the impact of A. tumefaciens on plant signal transduction when using A. tumefaciens-mediated transient expression assays to investigate ABA-regulated processes or pathogenicity and plant defence mechanisms.
Conflict of interest statement
Figures
Similar articles
-
Antagonism between salicylic and abscisic acid reflects early host-pathogen conflict and moulds plant defence responses.Plant J. 2009 Aug;59(3):375-86. doi: 10.1111/j.1365-313X.2009.03875.x. Epub 2009 Apr 24. Plant J. 2009. PMID: 19392690
-
Agroinfiltration by cytokinin-producing Agrobacterium sp. strain GV3101 primes defense responses in Nicotiana tabacum.Mol Plant Microbe Interact. 2014 Nov;27(11):1175-85. doi: 10.1094/MPMI-04-14-0114-R. Mol Plant Microbe Interact. 2014. PMID: 25054409
-
Interplay between JA, SA and ABA signalling during basal and induced resistance against Pseudomonas syringae and Alternaria brassicicola.Plant J. 2008 Apr;54(1):81-92. doi: 10.1111/j.1365-313X.2007.03397.x. Epub 2007 Dec 15. Plant J. 2008. PMID: 18088307
-
Differential volatile emissions and salicylic acid levels from tobacco plants in response to different strains of Pseudomonas syringae.Planta. 2003 Sep;217(5):767-75. doi: 10.1007/s00425-003-1039-y. Epub 2003 Apr 24. Planta. 2003. PMID: 12712338
-
A novel role of PR2 in abscisic acid (ABA) mediated, pathogen-induced callose deposition in Arabidopsis thaliana.New Phytol. 2013 Dec;200(4):1187-99. doi: 10.1111/nph.12436. Epub 2013 Aug 19. New Phytol. 2013. PMID: 23952213
Cited by
-
Comparative Membrane-Associated Proteomics of Three Different Immune Reactions in Potato.Int J Mol Sci. 2018 Feb 10;19(2):538. doi: 10.3390/ijms19020538. Int J Mol Sci. 2018. PMID: 29439444 Free PMC article.
-
Agromonas: a rapid disease assay for Pseudomonas syringae growth in agroinfiltrated leaves.Plant J. 2021 Feb;105(3):831-840. doi: 10.1111/tpj.15056. Epub 2021 Jan 12. Plant J. 2021. PMID: 33124734 Free PMC article.
-
'Candidatus Phytoplasma mali' SAP11-Like protein modulates expression of genes involved in energy production, photosynthesis, and defense in Nicotiana occidentalis leaves.BMC Plant Biol. 2024 May 13;24(1):393. doi: 10.1186/s12870-024-05087-4. BMC Plant Biol. 2024. PMID: 38741080 Free PMC article.
-
Transient proteolysis reduction of Nicotiana benthamiana-produced CAP256 broadly neutralizing antibodies using CRISPR/Cas9.Front Plant Sci. 2022 Aug 18;13:953654. doi: 10.3389/fpls.2022.953654. eCollection 2022. Front Plant Sci. 2022. PMID: 36061808 Free PMC article.
-
Transcriptome Analysis of Plant Hormone-Related Tomato (Solanum lycopersicum) Genes in a Sunlight-Type Plant Factory.PLoS One. 2015 Dec 1;10(12):e0143412. doi: 10.1371/journal.pone.0143412. eCollection 2015. PLoS One. 2015. PMID: 26624004 Free PMC article.
References
-
- Chilton MD, Drummond MH, Merio DJ, Sciaky D, Montoya AL, et al. Stable incorporation of plasmid DNA into higher plant cells: the molecular basis of crown gall tumorigenesis. Cell. 1977;11:263–271. - PubMed
-
- Van Larebeke N, Engler G, Holsters M, Van den Elsacker S, Zaenen I, et al. Large plasmid in Agrobacterium tumefaciens essential for crown gall-inducing ability. Nature. 1974;252:169–170. - PubMed
-
- Hellens R, Mullineaux P, Klee H. A guide to Agrobacterium binary Ti vectors. Trends Plant Sci. 2000;5:446–451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
