

Alternative splicing and extensive RNA editing of human TPH2 transcripts
- PMID: 20126463
- PMCID: PMC2813293
- DOI: 10.1371/journal.pone.0008956
Alternative splicing and extensive RNA editing of human TPH2 transcripts
Abstract
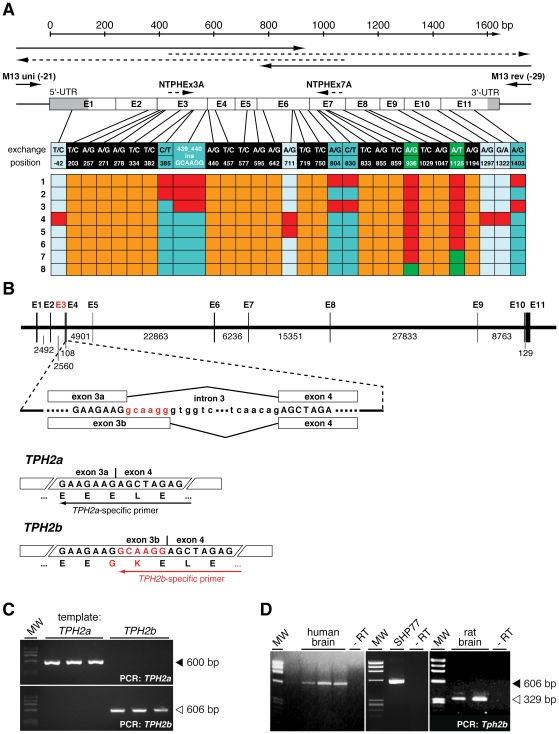
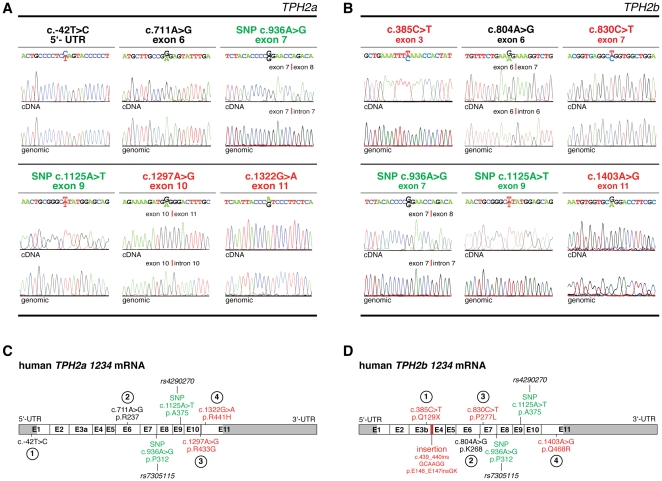
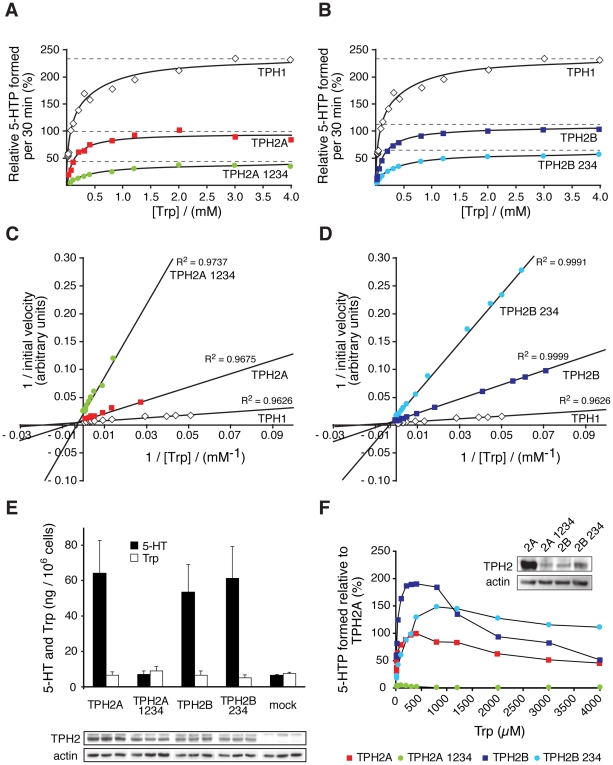
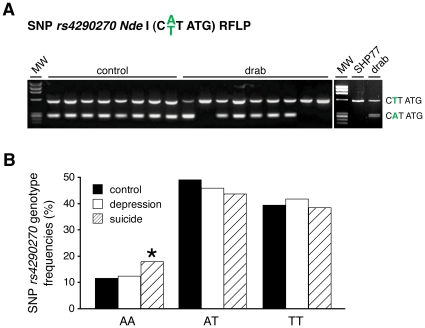
Brain serotonin (5-HT) neurotransmission plays a key role in the regulation of mood and has been implicated in a variety of neuropsychiatric conditions. Tryptophan hydroxylase (TPH) is the rate-limiting enzyme in the biosynthesis of 5-HT. Recently, we discovered a second TPH isoform (TPH2) in vertebrates, including man, which is predominantly expressed in brain, while the previously known TPH isoform (TPH1) is primarly a non-neuronal enzyme. Overwhelming evidence now points to TPH2 as a candidate gene for 5-HT-related psychiatric disorders. To assess the role of TPH2 gene variability in the etiology of psychiatric diseases we performed cDNA sequence analysis of TPH2 transcripts from human post mortem amygdala samples obtained from individuals with psychiatric disorders (drug abuse, schizophrenia, suicide) and controls. Here we show that TPH2 exists in two alternatively spliced variants in the coding region, denoted TPH2a and TPH2b. Moreover, we found evidence that the pre-mRNAs of both splice variants are dynamically RNA-edited in a mutually exclusive manner. Kinetic studies with cell lines expressing recombinant TPH2 variants revealed a higher activity of the novel TPH2B protein compared with the previously known TPH2A, whereas RNA editing was shown to inhibit the enzymatic activity of both TPH2 splice variants. Therefore, our results strongly suggest a complex fine-tuning of central nervous system 5-HT biosynthesis by TPH2 alternative splicing and RNA editing. Finally, we present molecular and large-scale linkage data evidencing that deregulated alternative splicing and RNA editing is involved in the etiology of psychiatric diseases, such as suicidal behaviour.
Conflict of interest statement
Figures
References
-
- Walther DJ, Bader M. A unique central tryptophan hydroxylase isoform. Biochemical Pharmacology. 2003;66:1673–1680. - PubMed
-
- Fitzpatrick PF. Tetrahydropterin-dependent amino acid hydroxylases. Annual Review of Biochemistry. 1999;68:355–381. - PubMed
-
- Rosen JB, Donley MP. Animal studies of amygdala function in fear and uncertainty: relevance to human research. Biological Psychiatry. 2006;73:49–60. - PubMed
-
- Baxter MG, Murray EA. The amygdala and reward. Nature Reviews Neuroscience. 2002;3:563–573. - PubMed
-
- Talarovicova A, Krskova L, Kiss A. Some assessments of the amygdala role in suprahypothalamic neuroendocrine regulation: a minireview. Endocrine Regulations. 2007;41:155–162. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
