

Axonal velocity distributions in neural field equations
- PMID: 20126532
- PMCID: PMC2813262
- DOI: 10.1371/journal.pcbi.1000653
Axonal velocity distributions in neural field equations
Abstract
By modelling the average activity of large neuronal populations, continuum mean field models (MFMs) have become an increasingly important theoretical tool for understanding the emergent activity of cortical tissue. In order to be computationally tractable, long-range propagation of activity in MFMs is often approximated with partial differential equations (PDEs). However, PDE approximations in current use correspond to underlying axonal velocity distributions incompatible with experimental measurements. In order to rectify this deficiency, we here introduce novel propagation PDEs that give rise to smooth unimodal distributions of axonal conduction velocities. We also argue that velocities estimated from fibre diameters in slice and from latency measurements, respectively, relate quite differently to such distributions, a significant point for any phenomenological description. Our PDEs are then successfully fit to fibre diameter data from human corpus callosum and rat subcortical white matter. This allows for the first time to simulate long-range conduction in the mammalian brain with realistic, convenient PDEs. Furthermore, the obtained results suggest that the propagation of activity in rat and human differs significantly beyond mere scaling. The dynamical consequences of our new formulation are investigated in the context of a well known neural field model. On the basis of Turing instability analyses, we conclude that pattern formation is more easily initiated using our more realistic propagator. By increasing characteristic conduction velocities, a smooth transition can occur from self-sustaining bulk oscillations to travelling waves of various wavelengths, which may influence axonal growth during development. Our analytic results are also corroborated numerically using simulations on a large spatial grid. Thus we provide here a comprehensive analysis of empirically constrained activity propagation in the context of MFMs, which will allow more realistic studies of mammalian brain activity in the future.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
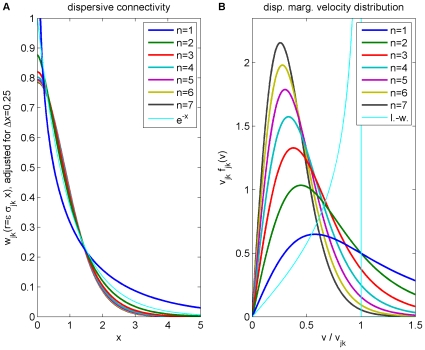
 for different powers
for different powers  , which has been adjusted to match an exponential decay (thin curve). While the curves are continuous here, adjustment with Eq. (25) assumes a bin size
, which has been adjusted to match an exponential decay (thin curve). While the curves are continuous here, adjustment with Eq. (25) assumes a bin size  , see text for details. (B) Marginal velocity distribution
, see text for details. (B) Marginal velocity distribution  for different powers
for different powers  . Note that concerning the dimensionless ratio
. Note that concerning the dimensionless ratio  one obtains
one obtains  . The long-wavelength approximation
. The long-wavelength approximation  of Eq. (36) is shown for comparison as thin curve. See Eqns. (19) and (32) for (A) and (B), respectively.
of Eq. (36) is shown for comparison as thin curve. See Eqns. (19) and (32) for (A) and (B), respectively.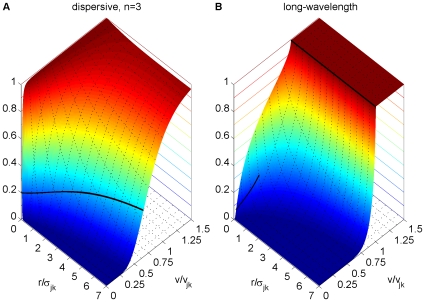
 as in Eq. (30). Dotted black lines on the base and on the plot surface show a grid of
as in Eq. (30). Dotted black lines on the base and on the plot surface show a grid of  and
and  values, solid black lines on the plot surface show the positions of the maxima of the unintegrated distributions. (A) Dispersive propagator for
values, solid black lines on the plot surface show the positions of the maxima of the unintegrated distributions. (A) Dispersive propagator for  , where
, where  corresponds to Eqn. (28). (B) Long-wavelength approximation, where
corresponds to Eqn. (28). (B) Long-wavelength approximation, where  integrates Eqn. (33). We set
integrates Eqn. (33). We set  and
and  for comparison.
for comparison.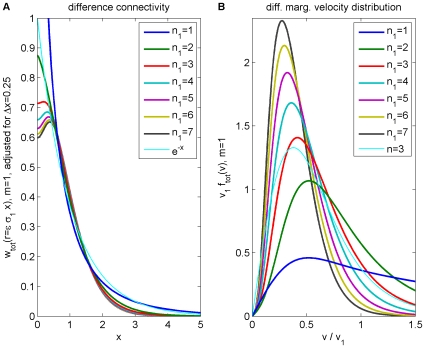
 . (A) Synaptic connectivity fit to an exponential decay (thin curve), Eqns. (47) and (49) are used. (B) Marginal velocity distribution Eq. (51). The dispersive
. (A) Synaptic connectivity fit to an exponential decay (thin curve), Eqns. (47) and (49) are used. (B) Marginal velocity distribution Eq. (51). The dispersive  case is shown as thin curve for comparison.
case is shown as thin curve for comparison.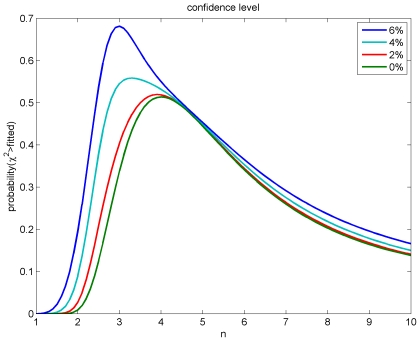
 of Eq. (54) was varied in steps of 0.1 for four different uncertainties of the observed threshold diameters
of Eq. (54) was varied in steps of 0.1 for four different uncertainties of the observed threshold diameters  . The assumed relative diameter error reflects mainly differential shrinkage. As confidence level the probability that
. The assumed relative diameter error reflects mainly differential shrinkage. As confidence level the probability that  is greater than the fitted
is greater than the fitted  is shown.
is shown.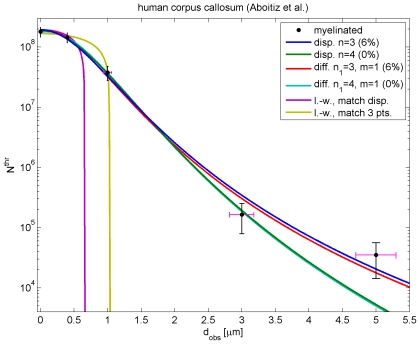
 and
and  fits are shown, which are optimal assuming
fits are shown, which are optimal assuming  equal to 6% and 0%, respectively. This relative diameter error (magenta error bars: 6%) reflects mainly differential shrinkage. Corresponding difference propagator fits are also shown, which have basically the same confidence levels. Thus these data cannot distinguish the dispersive and difference models, and the former is preferred for its computational simplicity. For the long-wavelength propagator a reasonable fit with Eq. (56) to all data cannot be obtained. Two curves are shown: one matching the median velocity of the dispersive
equal to 6% and 0%, respectively. This relative diameter error (magenta error bars: 6%) reflects mainly differential shrinkage. Corresponding difference propagator fits are also shown, which have basically the same confidence levels. Thus these data cannot distinguish the dispersive and difference models, and the former is preferred for its computational simplicity. For the long-wavelength propagator a reasonable fit with Eq. (56) to all data cannot be obtained. Two curves are shown: one matching the median velocity of the dispersive  case, the other fitting only the first three data points with
case, the other fitting only the first three data points with  .
.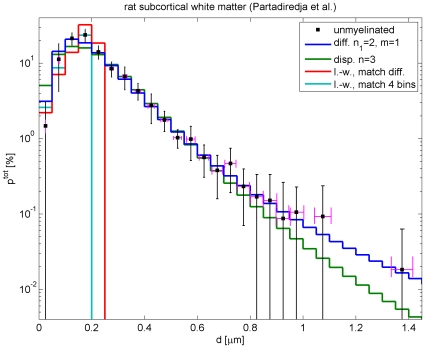
 (magenta error bars) has been assumed to reflect mostly differential shrinkage, but fit dependence on this is mild. Fit results using the difference Eq. (57), and its dispersive counterpart Eq. (58), are collected in Tab. 4. For unmyelinated axons the optimal fit with
(magenta error bars) has been assumed to reflect mostly differential shrinkage, but fit dependence on this is mild. Fit results using the difference Eq. (57), and its dispersive counterpart Eq. (58), are collected in Tab. 4. For unmyelinated axons the optimal fit with  ,
,  is shown. For comparison, the optimal
is shown. For comparison, the optimal  fit with the dispersive propagator is also displayed. It is viable, but has a three times larger
fit with the dispersive propagator is also displayed. It is viable, but has a three times larger  . For the long-wavelength propagator a reasonable fit with Eq. (59) to all data cannot be obtained. Two curves are shown for illustration: one matching the median velocity of the difference
. For the long-wavelength propagator a reasonable fit with Eq. (59) to all data cannot be obtained. Two curves are shown for illustration: one matching the median velocity of the difference  ,
,  case, the other fitting only the first four data points.
case, the other fitting only the first four data points.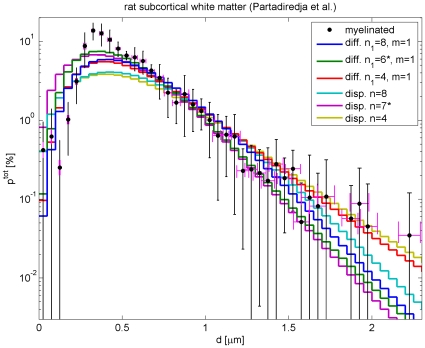
 and
and  , with
, with  in both cases. Systematic deviations from data around
in both cases. Systematic deviations from data around  are obvious, but fit quality remains tolerable with a confidence level of 53.45% for
are obvious, but fit quality remains tolerable with a confidence level of 53.45% for  . Even larger
. Even larger  can increase the confidence level to about 70%. For comparison, dispersive fits with orders
can increase the confidence level to about 70%. For comparison, dispersive fits with orders  and
and  are also shown. Their
are also shown. Their  is almost a factor two larger, rendering their confidence level negligible. Fits with the long-wavelength propagator are not show, but fail drastically, cf. Fig. 6. The curves marked with a
is almost a factor two larger, rendering their confidence level negligible. Fits with the long-wavelength propagator are not show, but fail drastically, cf. Fig. 6. The curves marked with a  show additional fits for diameters
show additional fits for diameters  only, i.e., without the first four data bins. Then one can find optimal fit orders for both propagators. These fits are of comparable, excellent quality compared to the reduced data set. But both predict too many small diameter fibres, and hence have negligible confidence levels compared to the full data set, with the dispersive
only, i.e., without the first four data bins. Then one can find optimal fit orders for both propagators. These fits are of comparable, excellent quality compared to the reduced data set. But both predict too many small diameter fibres, and hence have negligible confidence levels compared to the full data set, with the dispersive  again being about two times larger.
again being about two times larger.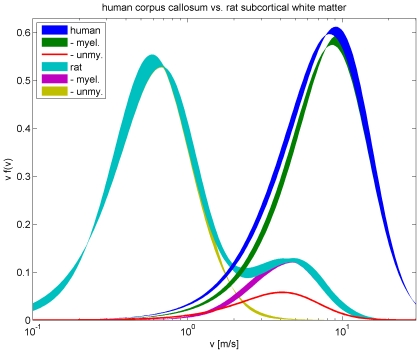
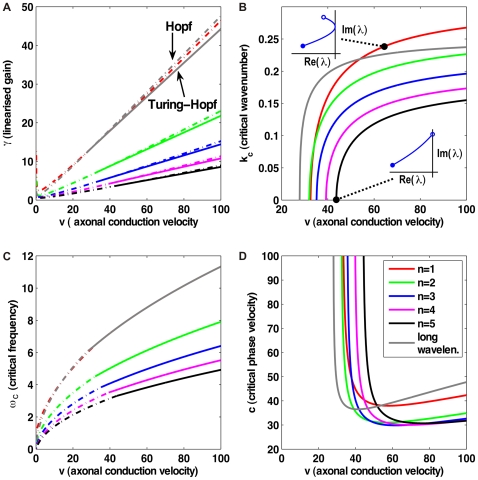
 and determining
and determining  ,
,  , and the critical linearized gain
, and the critical linearized gain  . All other model parameters remain at the values discussed in the text. (A) Solid curves represent Turing-Hopf bifurcations (
. All other model parameters remain at the values discussed in the text. (A) Solid curves represent Turing-Hopf bifurcations ( ), dot-dashed curves Hopf bifurcations (
), dot-dashed curves Hopf bifurcations ( ). Results for orders
). Results for orders  of the dispersive propagator and for the long-wavelength model are shown. Above the Turing-Hopf curves travelling waves emerge, whereas above the Hopf curves bulk oscillations are seen. Stability will be lost at a given
of the dispersive propagator and for the long-wavelength model are shown. Above the Turing-Hopf curves travelling waves emerge, whereas above the Hopf curves bulk oscillations are seen. Stability will be lost at a given  through the less stable bifurcation, which has smaller critical
through the less stable bifurcation, which has smaller critical  . (B) Critical wavenumber
. (B) Critical wavenumber  of the Turing-Hopf bifurcation. Insets show the position in the complex plane of the most weakly damped pole under variations of
of the Turing-Hopf bifurcation. Insets show the position in the complex plane of the most weakly damped pole under variations of  (open circles
(open circles  , closed circles
, closed circles  ) for the dispersive model at the indicated
) for the dispersive model at the indicated  . (C) Critical frequency
. (C) Critical frequency  of the less stable bifurcation. (D) Critical phase velocity
of the less stable bifurcation. (D) Critical phase velocity  , shown where Turing-Hopf is the less stable bifurcation.
, shown where Turing-Hopf is the less stable bifurcation.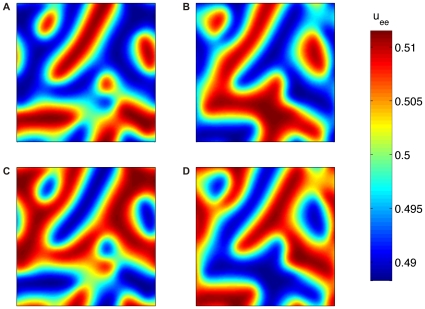
 spaced a quarter of the average temporal oscillation period apart. The dispersive propagator model of Eqns. (66)–(68) was computed for
spaced a quarter of the average temporal oscillation period apart. The dispersive propagator model of Eqns. (66)–(68) was computed for  and
and  with
with  chosen well beyond the Turing-Hopf critical value, cf. Fig. 9A. Spatial derivatives were approximated using finite differences on a regular square grid of
chosen well beyond the Turing-Hopf critical value, cf. Fig. 9A. Spatial derivatives were approximated using finite differences on a regular square grid of  with spacing
with spacing  . The resulting system of equations was rewritten as a first-order system and integrated using
. The resulting system of equations was rewritten as a first-order system and integrated using  . See also the supplementary Video S1 for the corresponding animation.
. See also the supplementary Video S1 for the corresponding animation.References
-
- Wilson HR, Cowan JD. A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue. Kybernetik. 1973;13:55–80. - PubMed
-
- Lopes da Silva FH, Hoeks A, Smits H, Zetterberg LH. Model of brain rhythmic activity: The alpha-rhythm of the thalamus. Kybernetik. 1973;15:27–37. - PubMed
-
- Freeman WJ. Mass action in the nervous system: Examination of the neurophysiological basis of adaptive behavior through the EEG. New York: Academic Press, 1st edition; 1975. Cf. http://sulcus.berkeley.edu/MANSWWW/MANSWWW.html.
-
- Lopes da Silva FH, van Rotterdam A, Barts P, van Heusden E, Burr W. Models of neuronal populations: The basic mechanism of rhythmicity. Prog Brain Res. 1975;45:281–308. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
